Dythundrakët
Dythundrakët (Artiodaktilat, nga greqishtja e lashtë ἄρτιος , ártios , që do të thotë 'madje', dhe δάκτυλος , dáktylos , do të thotë "gisht') janë kafshë thundrake - të cilat si thundra kanë vetëm dy gishtërijn. Tre gishtërinjtë e tjerë janë ose të pranishëm, në mungesë, në vestigial , ose duke treguar në anën e pasme. Në të kundërt, njëthundrakët mbajnë peshë në një (një numër të rastësishëm) nga pesë gishtërinjtë: gishtin e tretë. Një tjetër ndryshim midis të dyve është se thundra e barabartë me gishtin e tretë celulozën bimore në një ose duke pasur një stomakme katë dhoma sesa në zorrët e tyre siç bëjnë njëthundrakët.
| Dythundrakët | |
|---|---|

| |
| Dythundrakët. Ndërsa cetacët siç janë balenat dhe delfinët nuk janë artiodaktila normale, ato i përkasin kadadhës ; Artiodaktilat nganjëherë quhen Cetartiodaktila për të pasqyruar këtë. [1] | |
| Klasifikimi shkencor | |
| Mbretëria: | Animalia |
| Filumi: | Chordata |
| Klasa: | Mammalia |
| Magnorendi: | Boreoeutheria |
| Dega: | Laurasiatheria |
| Dega: | Scrotifera |
| Grandorder: | Ferungulata |
| Dega: | Ungulata |
| Rendi: | Artiodactyla Owen, 1848 |
| Subdivisions | |
Cetacët ujore (balenat , delfinët dhe porpoisët) evoluan nga dythundrakët, kështu që klasifikimi modern taksonomik ndërthur të dy nën emrin Cetartiodaktila.
Përafërsisht janë 270 specie toksore duke përfshir derrat , pekarët , hipopotamët , antilopat ,drerët-minj , drerët , gjirafat , devetë , lamat , alpakat , delet , dhitë dhe lopët . Shumë nga këto kanë një rëndësi të madhe dietike, ekonomike dhe kulturore për njerëzit.
Evolucion
RedaktoFosilet më të vjetra të dythundrakëve datojnë që nga Eoceni i hershëm (rreth 53 milion vjet më parë). Meqenëse këto gjetje pothuajse njëkohësisht u shfaqën në Evropë, Azi dhe Amerikën e Veriut, është shumë e vështirë të përcaktohet me saktësi origjina e artiodaktilëve. Fosilet klasifikohen se i përkasin familjes Dichobunidae ; anëtari i tyre më i njohur dhe më i ruajtur është Diacodexis .[2] Këto ishin kafshë të vogla, disa të vogla si lepujt , me një ndërtim të paktë, këmbë të holla dhe një bisht të gjatë. Këmbët e tyre të pasme ishin shumë më të gjata se këmbët e tyre të përparme. Fillim të Eocene mesme pa shfaqjen e paraardhësit e më të madhe të gjitarëve të sotëm.[3]

Dy familje dikur të përhapura, por tani të zhdukura,të dythundrakëve ishin Enteledontidae dhe Anthracotheriidae . Entelodontët ekzistonin nga Eoceni i mesëm deri në Miocenin e hershëm në Euroazi dhe Amerikën e Veriut. Kishin një trup të trashë me këmbë të shkurtra dhe kokë masive, e cila karakterizohej nga dy gunga në kockën e nofullës së poshtme. Anthracotherët kishte një ndërtim të madh, si derriat, me këmbë të shkurtra dhe një surrat të zgjatur . Ky grup u shfaq në Eocenë e mesme deri në Pliocen, dhe u përhap në të gjithë Eurazinë, Afrikën dhe Amerikën e Veriut. Anthracotherët mendohet se janë paraardhësit e hipopotamëve, dhe, në të njëjtën mënyrë, me siguri udhëhequr një mënyrë jetese të ngjashme ujore. Hipopotamët u shfaqën në Miocenin e vonë dhe pushtuan Afrikën dhe Azinë - ata kurrë nuk arritën në Amerikë.[3]
Devetë ( Tylopoda ) ishin, gjatë pjesësmë të madhe të Kenozoikut , të kufizuara në Amerikën e Veriut; forma të hershme si Cainotheriidae pushtuan Evropën. Ndër devetë e Amerikës së Veriut ishin grupe si Merycoidodontidae me këmbë të shkurtra . Ata së pari u shfaqën në Eocenin e vonë dhe zhvilluan një larmi të madhe të specieve në Amerikën e Veriut. Vetëm në Miocenin e vonë ose në Pliocenin e hershme ata migruan nga Amerika e Veriut në Euroazi.
Suina (përfshirë derrat ) kanë qenë që nga Eoceni. Në Eocenin e vonë ose Oligocen , dy familje qëndruan në Euroazi dhe Afrikë; pekarët , të cilët u zhdukën në Botën e Vjetër , ekzistojnë sot vetëm në Amerikë .

Dythundrtakët e Amerikës Jugore u zhvendosën vetëm në Pliocen, pasi ura tokësore në Isthmus e Panamasë u formua rreth tre milion vjet më parë. Me vetëm pekarët, lamoidët (ose lamat ), dhe lloje të ndryshme të drerëve capreoline , Amerika e Jugut ka relativisht më pak pjestar të artiodaktilave sesa kontinentet e tjera, përveç Australisë, e cila nuk ka specie vendase.
Taksonomia dhe filogjinia
Redakto
Klasifikimi i artiodaktilave u debatua ashpër sepse cetacët që banonin në oqean evoluan nga dythundrakët tokësor. Disa dythundrakë semiakuatik ( hipopotamët ) janë më të lidhura me cetacët që banojnë në oqean sesa me dythundrakët e tjerë.
Kjo e bën Artiodaktilën siç përcaktohet tradicionalisht një takson parafillete , pasi përfshin kafshët e prejardhura nga një paraardhës i zakonshëm , por nuk përfshin të gjithë pasardhësit e saj. Klasifikimi filogjenetik njeh vetëm taksat monofleike ; domethënë, grupe që vijnë nga një paraardhës i zakonshëm dhe përfshijnë të gjithë pasardhësit e tij. Për të adresuar këtë problem, rendi tradicional Artiodaktila dhe infrarendi Cetacea nganjëherë përfshihen në taksonin më përfshirës të Cetartiodaktila.[1] Një qasje alternative është të përfshini të dy,dythundrakët tokësor dhe cetacët që banojnë në oqean në një taksion të rishikuar Artiodaktila. [3]
Klasifikimi
Redakto- Rendi Artiodaktila/Klade Cetartiodaktila[3][4]
- Nënrendi Tylopoda
- Familja †Anoplotheriidae?
- Familja †Cainotheriidae
- Familja †Merycoidodontidae
- Familja †Agriochoeridae
- Familja Kamelidae: devetë dhe lamat (7 të gjalla dhe 13 lloje të zhdukura)
- Familja †Oromerycidae
- Familja †Xiphodontidae
- Klada Artiofabula
- Nënrendi Suina
- Familja Suidae: derrat (19 lloje)
- Familja Tayassuidae: pekarët (4 lloje)
- Familja †Sanitheriidae
- Klada Cetruminantia
- Klada Cetancodontamorpha
- Gjinia †Andrewsarchus?
- Familja †Entelodontidae
- Nënrendi Whippomorpha
- Familja †Raoellidae
- Nënrendi Dichobunoidea – parafiletik e Cetacea dhe Raoellidae
- Familja †Dichobunidae
- Familja †Helohyidae
- Familja †Choeropotamidae
- Familja †Cebochoeridae
- Familja †Mixtotheriidae
- Infrarendi Ancodonta
- Familja †Anthracotheriidae – parafiletik me Hippopotamidae
- Familja Hippopotamidae: hipopotamët (dy lloje)
- Infrarendi Cetacea: balenat (rreth 90 lloje)
- Parvrendi †Archaeoceti
- Familja †Pakicetidae
- Familja †Ambulocetidae
- Familja †Remingtonocetidae
- Familja †Basilosauridae
- Parvrendi Mysticeti: balenat me mustaqe
- Mbifamilja Balaenoidea: balenat e duhura
- Familja Balaenidae: (katë lloje)
- Familja Cetotheriidae: balena e duhur e vogël (one species)
- Mbifamija Balaenopteroidea: balenat me hulli
- Familja Balaenopteridae: balenat e mëdha dhe gungace (tetë lloje)
- Familja Eschrichtiidae: balena e hirtë (një lloj)
- Mbifamilja Balaenoidea: balenat e duhura
- Parvrendi Odontoceti: balenat me dhëmbë
- Mbifamilja Delphinoidea: delfinët oqeanik,porpisët
- Familja Delphinidae: delfinët e vërtet oqeanik (38 lloje)
- Familja Monodontidae: balenat artike; narvhali dhe beluga (dy lloje)
- Familja Phocoenidae: porpoisët (gjashtë lloje)
- Mbifamija Physeteroidea: kashalota
- Familja Kogiidae: kashalota e vogël (dy lloje)
- Familja Physeteridae: kashalota (një lloj)
- Mbifamilja Platanistoidea: river dolphins
- Familja Iniidae: delfinët e lumejnve amerikanojugor (dy lloje)
- Familja Lipotidae: delfini i lumit kinez (nj) lloj,ndoshta i zhdukur
- Familja Platanistidae: delfini i lumejnve aziatikojugor (one species)
- Familja Pontoporiidae: delfini i La Platas (one species)
- Mbifamilja Ziphioidea
- Familja Ziphidae: balenat me sqep (22 lloje)
- Mbifamilja Delphinoidea: delfinët oqeanik,porpisët
- Parvrendi †Archaeoceti
- Klada Ruminantiamorpha
- Nënrendi Ruminantia
- Infrarendi Tragulina
- Familja †Amphimerycidae
- Familja †Prodremotheriidae
- Familja †Protoceratidae
- Familja †Hypertragulidae
- Familja †Praetragulidae
- Familja Tragulidae: çevrotainët (dhjetë lloje)
- Familja †Archaeomerycidae
- Familja †Lophiomerycidae
- Infrarendi Pecora
- Familja †Gelocidae
- Familja †Palaeomerycidae
- Familja Antilocapridae: pronghorni (një lloj)
- Familja †Climacoceratidae
- Familja Girafidae: okapi dhe katër lloje të gjirafave (pesë lloje në total)
- Familja †Hoplitomerycidae
- Familja Cervidae: drerët (49 lloje)
- Familja †Leptomerycidae
- Familja Moshidae: drerët e myshqeve (shtatë lloje)
- Familja Bovidae: lopët, buallicat, dhitë, antelopat, dhe të tjerët (135 species)
- Infrarendi Tragulina
- Nënrendi Ruminantia
- Klada Cetancodontamorpha
- Nënrendi Suina
- Nënrendi Tylopoda
Historia e hulumtimit
Redakto
Në vitet 1990, sistematika biologjike përdori jo vetëm morfologjinë dhe fosilet për të klasifikuar organizmat, por edhe biologjinë molekulare . Biologjia molekulare përfshin renditjen e ADN-së dhe ARN-së së një organizmi dhe krahasimin e sekuencës me atë të qenieve të tjera të gjalla - sa më të ngjashëm të jenë, aq më të afërt janë të lidhur. Krahasimi i materialit gjenetik të papërpunuar dhe cetacët të barabartë ka treguar se të afërmit më të afërt të gjallë të balenave dhe hipopotamëve është grupi parafitletik Artiodaktila.
Dan Graur dhe Desmond Higgins ishin ndër të parët që arritën në këtë përfundim dhe përfshinë një punim të botuar në 1994.[5] Sidoqoftë, ata nuk i njohën hipopotamët dhe i klasifikuan ripërtypësit si grupi motër i cetacëve. Studimet pasuese vërtetuan lidhjen e ngushtë midis hipopotamëve dhe cetacëve; këto studime u bazuan në gjenet e kazeinës ,[6] SINE ,[7] sekuenca fibrinogjene ,[8] sekuencat citokromi dhe ARN ,[1][9] IRBP (dhe vWF ) sekuencat e gjeneve, [10] receptorët adrenergjik ,[11] dhe apolipoproteinat .[12]
Në 2001, gjymtyrët fosile të një Pakicetus ( cetac amfibiod me madhësin e një ujku) dhe Ichthyolestes (një balenë e hershme me madhësinë e një dhelpre) u gjetën në Pakistan. Ata të dy ishin arkeocetë ("balenat antike") nga rreth 48 milion vjet më parë (në Eocen). Këto gjetje treguan se arkeocetët ishin më tokësore sesa mendohej më parë, dhe se ndërtimi special i talusit (kockës së këmbës) me një sipërfaqe të përbashkët të mbështjellë të dyfishtë, që më parë mendohej se ishte unik për dythundrakët dhe cetacët e hershme.[13] Mesonychidët, një lloj tjetër i dythundrakëve, nuk e tregoi këtë ndërtim të veçantë të talusit, dhe kështu u konstatua se nuk kishin të njëjtët paraardhës si cetacët.

Cetacët më të vjetër datojnë që nga Eoceni i hershëm (53 milion vjet më parë), ndërsa hipopotami më i vjetër i njohur daton vetëm në Miocen (15 milion vjet më parë). Disa dyshime janë lindur në lidhje me marrëdhëniet midis të dyve, pasi ekziston një hendek prej 40 milion vitesh midis paraqitjeve të tyre të para në procesverbalin fosil. Duket se nuk ka të ngjarë që të ketë hipopotami stërgjyshor që nuk kanë mbetur mbetje, duke pasur parasysh numrin e madh të fosileve të humbura. Disa studime propozuan që shfaqja e vonë e hipopotamëve është sepse ata janë të afërm të sekondareve dhe të ndarë kohët e fundit, por gjetjet molekulare bien në kundërshtim me këtë. Prandaj, hulumtimi është përqendruar me anthrakortherët (familja Anthracotheriidae); një që daton nga Eoceni në Miocene u shpall të jetë "si-hipopotam" pas zbulimit në shekullin e 19-të. Sidoqoftë besohej se cetacët dhe anthrakothererët zbritnin nga një paraardhës i zakonshëm, dhe se hipopotamët u zhvilluan nga antrakoterët. Një studim i publikuar në 2015 ishte në gjendje ta konfirmonte këtë, por gjithashtu zbuloi se hipopotamët rridhnin nga antracoterapët më të vjetër.[9][14] Gjinia Epirigenys nga Afrika Lindore është grupi motër i hipopotamëve.
Klasifikimi morfologjik i Artiodaktila
RedaktoLinnaeus postuloi një lidhje të ngushtë midis deveve dhe ripërtypësve që nga mesi i viteve 1700. Henri de Blainville njohu anatominë e ngjashme të gjymtyrëve të derrave dhe hipopotamëve, dhe zoologu britanik Richard Owen krijuan termin "dythundrakët" dhe emrin shkencor "Artiodaktila" në 1848.[nevojitet citimi]
Morfologjia e brendshme (kryesisht stomaku dhe molarët) janë përdorur për klasifikim. Suitat (përfshirë derrat ) dhe hipopotamët kanë molarë me rrënjë të zhvilluara mirë dhe stomak të thjeshtë që tret ushqimin. Kështu, ata u grupuan së bashku si jo-ripërtypës (Porkine). Të gjithë dythundrakët kanë një molarë me një konstruksion selenodont ( kreshta në formë gjysmëhënës) dhe kanë aftësinë të piqen , gjë që kërkon rregullimin e ushqimit dhe ripërtypësit e tij. Dallimet në ndërtimin e stomakut treguan se shpërthimi evoluoi në mënyrë të pavarur midis tylopodëve dhe ripërtypësve ; prandaj, tylopodët u përjashtuan nga Ruminantia .
Taksonomia që u pranua gjerësisht nga fundi i shekullit të 20 ishte:[15][nevojitet citimi i plotë]
Even-toed ungulates
|
| ||||||||||||||||||||||||
Klasifikimi morfologjik i Cetacea
Redakto
Cetacët moderne janë krijesa deti të përshtatura, të cilat, morfologjike, kanë pak të përbashkëta me gjitarët tokësor; ato janë të ngjashme me gjitarët e tjerë detarë , siç janë fokat dhe lopët e detit , për shkak të evolucionit konvergjent . Sidoqoftë, ata evoluan nga gjitarët fillimisht tokësorë. Paraardhësit më të mundshëm mendohej se ishin mesonychidët - kafshë të mëdha, mishngrënëse nga Kenozoiku i hershëm ( Paleoceni dhe Eoceni), të cilat kishin thundra në vend të kthetrave në këmbë. Molarët e tyre ishin përshtatur në një dietë mishngrënëse, duke u ngjarë me dhëmbët e balenave moderne me dhëmbë , dhe, ndryshe nga gjitarët e tjerë, kanë një ndërtim uniform. [nevojitet citimi]
Marrëdhëniet e dyshuara mund të tregohen si më poshtë:[14][16]
| Paraxonia |
| ||||||||||||
Sistematika e brendshme
RedaktoGjetjet molekulare dhe indikacionet morfologjike sugjerojnë që artiodaktilët siç përcaktohen tradicionalisht janë parafiletë në lidhje me cetacët. Cetacët janë fole thellësisht brenda të parës; të dy grupet së bashku formojnë një takson monofiletik, për të cilën emri Cetartiodaktila është përdorur ndonjëherë. Nomenklatura moderne ndan Artiodaktila (ose Cetartiodaktila) në katër taksa vartëse: devet (Tylopoda), derra dhe pekarët (Suina), ripërtypësit (Ruminantia), dhe hipopotamët plus balenat (Whipomorfa).
Linjat e supozuara brenda Artiodaktila mund të përfaqësohen në kladogramin vijues : [17][18][19][20][21]
| Artiodactyla |
| ||||||||||||||||||||||||||||||


Katër taksat e përmbledhura të Artiodaktila ndahen në dhjetë familje të gjalla:
The four summarized Artiodactyla taxa are divided into ten extant families:[22]
- Kamelidët ( Tylopoda ) përbëjnë vetëm një familje, Kamelidae . Ёshtë një nënrend i varfër me speciet me origjinë nga Amerika veriore [23] që është përshtatur mirë me habitatet ekstreme - devet dromedar dhe bakterian në shkretëtirat e Botës së Vjetër dhe guanakot , lamat, vikunjat , dhe alpakas në rajonet e larta malore të Amerikës së Jugut.
- Krijesat si derri ( Suina ) përbëhen nga dy familje:
- Derrat ( Suidae ) janë të kufizuar në Botën e Vjetër. Këto përfshijnë derrin e egër dhe formën shtëpiake, derrin shtëpiak .
- Pekarët ( Taiasuidae ) janë emëruar pas gjëndrave në bark dhe janë indigjenë për Amerikën Qendrore dhe Jug.
- Ruminantët ( Ruminantia ) përbëhen nga gjashtë familje:
- Dreri-mi ( Tragulidae ) janë ripërtypësit më të vegjël dhe më primitivë; ata banojnë në pyjet e Afrikës dhe Azisë.
- Krijesat si gjirafat ( Girafidae ) janë të përbëra nga dy specie: gjirafat dhe okapi .
- Drerët e myshqeve ( Moshidae ) është indigjen për Azinë Lindore.
- Antilokapridët ( Antilocapridae ) të Amerikës së Veriut përmbajnë vetëm një specie të gjallë: pronghornin .
- Drerët ( Cervidae ) është i përbërë nga rreth 45 lloje, të cilat karakterizohen nga një çift brirësh (zakonisht vetëm te meshkujt). Ato janë përhapur nëpër Evropë, Azi dhe Amerikë. Ky grup përfshin, drerin e kuq , dreri brilopat , wapiti, dhe renët (karibut).
- Gjethët ( Bovidae ) janë më të pasura me specie. Midis tyre janë bagëtitë , delet , dhitë dhe antilopat .
- Whipomorfanët përfshijnë hipopotamët dhe cetacët:
- Hipopotamët ( Hipopotamidae ) përbëjnë dy grupe, hipopotamët dhe hipopotamët xhuxh .
- Balenat ( Cetacea ) përmbajnë 72 specie dhe dy parvrend: balenat me dhëmbë ( Odontoceti ) dhe balenat me mustaqe ( Mysticeti )
Megjithëse drerët, drerët e myshqeve dhe pronghornët janë përmbledhur tradicionalisht si qafat cervidët (Cervioidea), studimet molekulare japin rezultate të ndryshme - dhe jo-konsistente, kështu që çështja e sistematikës filogjenetike të infeksionit Pekora (ripërtypësit me brirë) për momentin, nuk mund të jetë u përgjigj.

Anatomia
Redakto
Artiodaktilët në përgjithësi janë katërkëmbësha . Janë të njohura dy tipe kryesore trupore: Suinidët dhe hipopotamëtt karakterizohen nga një trup i stukuar, këmbë të shkurtra dhe një kokë të madhe; devetë dhe ripërtypësit, sidoqoftë, kanë këmbë më të holla të ndërtuara dhe të dobëta. Madhësia ndryshon në mënyrë të konsiderueshme; anëtari më i vogël, dreri-mi, shpesh arrin një gjatësi trupi prej vetëm 45 cm (18 in) dhe një peshë prej 1.5 kilogramësh (3.3 lb). Anëtari më i madh, hipopotami, mund të rritet deri në 5 metra (16 ft) në gjatësi dhe të peshojë 4,5 tonë metrikë (5.0 ton të shkurtër), dhe gjirafa mund të rritet të jetë 5.5 metra (18 ft) dhe 4,7 metra (15 ft ) në gjatësinë e trupit. Të gjithë dythundrakët shfaqin një formë të dimorfizmit seksual : meshkujt janë vazhdimisht më të mëdhenj dhe më të rëndë se femrat.Tek drerët, vetëm meshkujt kanë brirë , dhe brirët e bovidëve janë zakonisht të vogla ose jo të pranishme tek femrat. Antelopat indiane mashkuj kanë një pallto shumë më të errët sesa femrat.
Pothuajse të gjithë dythundrakët kanë gëzof, përjashtim bën hipopotami gati pa flokë. Gëzofi ndryshon në gjatësi dhe ngjyrosje në varësi të habitatit. Speciet në rajone më të freskëta mund të kenë pak gëzof pallton e tyre. Palltot e kamufluara vijnë me ngjyra të toneve të verdha, gri, kafe ose të zeza.
Gjymtyrët
Redakto
Dythundrakët e mbajnë emrin e tyre sepse kanë një numër të barabartë gishtërinjsh (dy ose katër) - në disa pekar, këmbët e pasme kanë një ulje të numrit të gishtërinjve në tre. Aksi qendror i këmbës është midis shputës së tretë dhe të katërt. Gishti i parë i mungon tek artiodaktilët moderne, dhe mund të gjendet vetëm në gjenet e zhdukur tani. Gishtat e dytë dhe të pestë janë përshtatur ndryshe midis specieve:
-
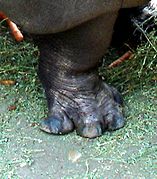 Hipopotamët i kanë të theksuar të katër gishtërinjtë
Hipopotamët i kanë të theksuar të katër gishtërinjtë -
 Për derrat dhe biungulatët e tjera gishtërinjtë e dytë dhe të pestë drejtohen prapa
Për derrat dhe biungulatët e tjera gishtërinjtë e dytë dhe të pestë drejtohen prapa -
 Kur devetë kanë vetëm dy gishtërinj, kthetrat shndërrohen në thonj.
Kur devetë kanë vetëm dy gishtërinj, kthetrat shndërrohen në thonj.



Kur devetë kanë vetëm dy gishtërinj të prerë, kthetrat shndërrohen në thonj (ndërsa të dyja janë bërë me keratin , kthetrat janë të lakuara dhe të theksuara ndërsa thonjtë janë të sheshtë dhe të shurdhër).[24] Këto kthetra përbëhen nga tre pjesë: pllaka (e sipërme dhe anët), e vetmja (fundi) dhe balte (e pasme). Në përgjithësi, kthetrat e këmbëve të përparme janë më të gjera dhe të zbardhura se ato të këmbëve të pasme, dhe gryka është larg më larg. Përveç deveve, të gjithë thundrakët gjithashtu vendosin majën e faranxhës më të madhe në tokë. [25]

Tek dythundrakët, eshtrat e stilopodiumit (kocka e krahut të sipërm ose kofshë) dhe zigopodiumet (tibia dhe fibula) zakonisht zgjaten. Muskujt e gjymtyrëve janë kryesisht të lokalizuara, gjë që siguron që artiodaktilët shpesh të kenë këmbë shumë të hollë. Një klavikul nuk është asnjëherë e pranishme, dhe skapula është shumë e shkathët dhe lëkundet përpara dhe mbrapa për lëvizje të shtuar kur vrapon. Ndërtimi i veçantë i këmbëve bën që këmbët të mos jenë në gjendje të rrotullohen, gjë që lejon një stabilitet më të madh kur vraponi me shpejtësi të mëdha. Për më tepër, shumë artiodaktilë më të vegjël kanë një trup shumë fleksibël, duke kontribuar në shpejtësinë e tyre duke rritur gjatësinë e tyre të përparimit.
Kafka
RedaktoShumë dythundrakë kanë një kokë relativisht të madhe. Kafka është e zgjatur dhe mjaft e ngushtë; kockat frontale është zgjeruar në afërsi të pasme dhe zhvendos të kockat parietal , e cila formon vetëm një pjesë të anën e kafkë (sidomos tek ripërtypësit).
Brirët
Redakto
Katër familje të dythundrakëve kanë një shtojcë kraniale. Këto Pekora , (me përjashtim të drerit të myshkut ), kanë një nga katër llojet e shtojcave kraniale: brirët e vërtetë, brirët e drerëve , osiconets ose pronghorn .[26]
Brirët e vërtetë kanë një bërthamë kockore që është e mbuluar në një copë të përhershme të keratinës, dhe gjenden vetëm në gjedhët . Brirët e drerëve janë struktura kockore që derdhen dhe zëvendësohen çdo vit; ato gjenden tek drerët (anëtarët e familjes Cervidae ). Ata rriten nga një dalje e përhershme e kockës ballore të quajtur pedikul dhe mund të degëzohen, si tek dreri bishtbardhë ( Odocoileus virginianus ), ose dreri brilopat ( Alces alces ). Osiconet janë struktura të përhershme kockash që bashkohen në kockat frontale ose parietale gjatë jetës së një kafshe dhe gjenden vetëm tek Girafidae . Pronghorn, ndërsa janë të ngjashëm me brirët në atë se ato kanë qepje keratinoze që mbulojnë bërthama të përhershme kockash, janë gjetherënëse.[27]
Të gjitha këto shtojca të burrrave mund të shërbejnë për sjellje, përleshje për privilegjin e çiftëzimit dhe mbrojtjen. Pothuajse në të gjitha rastet, ato janë seksuale me dimensione seksuale, dhe shpesh gjenden vetëm tek meshkujt.
Dhëmbët
Redakto
| Formula dentale | I | C | P | M | |
|---|---|---|---|---|---|
| 30–44 | = | 0–3 | 0–1 | 2–4 | 3 |
| 1–3 | 1 | 2–4 | 3 | ||
Ekzistojnë dy tendenca në aspektin e dhëmbëve brenda Artiodaktila. Suina dhe hipopotamët kanë një numër relativisht të madh të dhëmbëve (me disa derra që kanë 44); stomatologjia e tyre është më e përshtatur me një manipulim shtrydhës , i cili është karakteristik për të gjithë gjithçkangrënësit . Devetë dhe ripërtypësit kanë më pak dhëmbë; shpesh ekziston një diastemë e yawning , një hendek i caktuar tek dhëmbët, ku molarët janë të vendosur për shtypjen e lëndës bimore.
Incisorët janë ulur shpesh tek ripërtypësit, dhe janë plotësisht mungon në nofullën e sipërme . Kanunet janë të zmadhuara dhe të ngjashme me ato të Suina, dhe përdoren për gërmime në tokë dhe për mbrojtje. Tek ripërtypësit, dhëmbët e sipërme të meshkujve zgjerohen dhe përdoren si armë në specie të caktuara (drerët-minj, drerët e myshqeve, drerët e ujit ); speciet me brirë frontale zakonisht u mungojnë dhëmbët e sipërme. Kanunat e poshtëm të ripërtypësve ngjajnë me incizuesit, në mënyrë që këto kafshë të kenë tetë dhëmbë uniformë në pjesën ballore të nofullës së poshtme .
Molarët e derrit kanë vetëm disa gunga. Në të kundërt, devetë dhe ripërtypësit kanë gunga të cilat janë formë kreshta në formë gjysmëhënës ( selenodont ).
Shqisat
RedaktoArtiodaktilat kanë një sens të zhvilluar mirë të erës dhe ndjenjës së dëgjimit. Për dallim nga shumë gjitarë të tjerë, ata kanë një sens të dobët të shikimit - objektet lëvizëse janë shumë më të lehta për tu parë sesa ato të palëvizshme. Ngjashëm me shumë kafshë të tjera pre, sytë e tyre janë në anët e kokës, duke u dhënë atyre një pamje pothuajse panoramike.
Sistemi ushqimor
RedaktoRipërtypësit (Ruminantia dhe Tylopoda) e ripërtërijnë ushqimin e tyre ata derdh mbrapa dhe ripërtyp atë. Gojët e ruminantëve shpesh kanë gjëndra shtesë të pështymës , dhe mukoza me gojë shpesh quhet rëndë për të shmangur dëmtimin nga pjesët e bimëve të vështirë dhe për të lejuar transportimin më të lehtë të ushqimeve të përtypura afërsisht. Stomakët e tyre ndahen në tre deri në katër seksione: bajamja , retikuli , omasum dhe aboma .[28] Pasi ushqimi të hahet, ai përzihet me pështymë në gjellë dhe retikul dhe ndahet në shtresa të materialit të ngurtë kundrejt lëngut. Solidët grumbullohen së bashku për të formuar një bolus (i njohur edhe si kudër ); kjo rregullohet nga kontraktimet retikulare ndërsa glota është e mbyllur. Kur bolusi hyn në gojë, lëngu shtrydhet me gjuhën dhe përsëri gëlltitet. Bolusi përtypet ngadalë për ta përzier plotësisht me pështymë dhe për ta prishur atë. Ushqimi i gëlltitur kalon në "dhomën e fermentimit" (thashetheme dhe retikul), ku mbahet në lëvizje të vazhdueshme nga kontraktimet ritmike. Mikrobet celulitike ( bakteret , protozoa dhe kërpudhat ) prodhojnë celulazë , e cila është e nevojshme për të prishur celulozën që gjendet në materialin bimor. Kjo formë e tretjes ka dy avantazhe: bimët që nuk mund të dëmtohen nga speciet e tjera mund të treten dhe përdoren, dhe kohëzgjatja e konsumit aktual të ushqimit të shkurtohet; kafsha kalon vetëm një kohë të shkurtër jashtë në hapësirë me kokën në tokë - zhurmimi mund të bëhet më vonë, në një zonë të strehuar.[29]
Tylopoda (devet, lamat dhe alpakat) dhe çevrotainët kanë stomakë me tre koka, ndërsa pjesa tjetër e Ruminantia kanë stomakë me katër koka. Handikapi i një sistemi të rëndë tretës ka rritur presionin selektiv drejt gjymtyrëve që i lejojnë kafshës të shpëtojë shpejt nga grabitqarët. Shumica e specieve brenda Suinëve kanë një stomak të thjeshtë me dy koka që lejon një dietë gjithçkangrënëse. Babirusa , megjithatë, është një herbivor, dhe ka ekstra dhëmbët nofullës për të lejuar për përçapje e duhur të materialit bimor. Pjesa më e madhe e fermentimit ndodh me ndihmën e mikroorganizmave celulolitikë brenda cezumit të zorrëve të mëdha . Pekariët kanë një stomak kompleks që përmban katër ndarje. Stomaku i tyre i parë ka fermentim të realizuar nga mikrobet dhe ka nivele të larta të acidit yndyror të paqëndrueshëm ; është propozuar që stomaku i tyre i ndërlikuar i përparuar është një mjet për të ngadalësuar kalimin e tretjes dhe për të rritur efikasitetin e tretjes. Hipopotamët kanë stomakë me tre koka dhe nuk përtrihen. Konsumojnë rreth 68 kilogramë (150 lb) bar dhe lëndë të tjera bimore çdo natë. Ata mund të mbulojnë distanca deri në 32 kilometra (20 mi) për të marrë ushqim, të cilin ata e tretin me ndihmën e mikrobeve që prodhojnë celulazë. Të afërmit e tyre më të afërt që jetojnë, balenat, janë mishngrënës të detyrueshëm .
Për dallim nga dythundrakët e tjerë, derrat kanë një stomak të thjeshtë në formë thesi .[28] Disa artiodaktil, të tillë si dreri bishtbardhë , nuk kanë një fshikëz tëmthi. [30]
-
 Derrat (siç është ky derr savane ) kanë një stomak të thjeshtë në formë thesi.
Derrat (siç është ky derr savane ) kanë një stomak të thjeshtë në formë thesi. -
 Si me të gjithë ripërtypësit, dreri ka një stomak kaq të shumtë, i cili përdoret për tretjen më të mirë të ushqimeve bimore.
Si me të gjithë ripërtypësit, dreri ka një stomak kaq të shumtë, i cili përdoret për tretjen më të mirë të ushqimeve bimore.


Sistemi riprodhues
Redakto
Peniset e dythundrakëve janë në formë S-je në pushim dhe shtrihen në një xhep nën lëkurën në bark. Cavevernosa e korporatës është vetëm pak e zhvilluar; dhe një mbledhje bën që kjo lakim të zgjasë, gjë që çon në një zgjatje, por jo të një trashje të penisit . Cetacët kanë penis të ngjashme. Në disa dythundrakë, penisi përmban një strukturë të quajtur proces uretral.[31][32][33]
Testikujt janë të vendosura në scrotum dhe kështu jashtë zgavrën e barkut . Të vezoret e shumë femrave të zbresë - si testikujt zbresin nga shumë gjitarët meshkuj - dhe janë të afërt me gji legenit në nivelin e katërt rruazë mesit . Mitra ka dy brirë ( Bicornis mitra ).[34]
Të tjera
RedaktoNumri i gjëndrave qumështore është i ndryshueshëm dhe lidhet, si të gjithë gjitarët, me madhësi pjellore . Derrat, të cilët kanë madhësinë më të madhe të pjellave të të gjithë thundrakëve, kanë dy rreshta dhëmbësh të rreshtuar nga sqetulla në zonën e ijëve. Sidoqoftë, në shumicën e rasteve, dythundrakët kanë vetëm një ose dy çifte dhëmbësh. Në disa specie, këto formojnë një udder në rajonin e ijëve.
Gjëndrat sekretore në lëkurë janë të pranishme tek të gjithë speciet dhe mund të vendosen në vende të ndryshme, siç janë në sy; prapa brirëve, qafës ose shpinës; në këmbë; ose në rajonin anal.
Jetesa
RedaktoGama dhe habitati
RedaktoArtiodaktilët janë vendas për pothuajse të gjitha pjesët e botës, me përjashtim të Oqeanisë dhe Antarktidës . Njerëzit kanë futur artiodaktil të ndryshme në të gjithë botën si kafshë gjuetie.[35] Artiodaktilët banojnë pothuajse në çdo habitat, nga pyjet tropikale të shiut dhe stepat në shkretëtirat dhe rajonet e larta malore . Biodiversiteti më i madh mbizotëron në habitatet e hapura siç janë livadhet dhe pyjet e hapura.
Sjellja sociale
Redakto
Sjellja sociale e thundrakëve ndryshon nga speciet në specie. Në përgjithësi, ekziston një tendencë për t'u bashkuar në grupe më të mëdha, por disa jetojnë vetëm ose në çifte. Llojet që jetojnë në grupe shpesh kanë një hierarki , si tek meshkujt, ashtu edhe nga femrat. Disa specie gjithashtu jetojnë në grupe harem , me një mashkull, disa femra dhe pasardhësit e tyre të zakonshëm. Në speciet e tjera, femrat dhe të miturit qëndrojnë së bashku, ndërsa meshkujt janë të vetmuar ose jetojnë në grupe beqarie dhe kërkojnë femra vetëm gjatë sezonit të çiftëzimit .
Shumë artiodaktil janë territorial dhe shënojnë territorin e tyre, për shembull, me sekrecione gjëndre ose urinë . Përveç specieve sedentare gjatë gjithë vitit, ka kafshë që migrojnë sezonalisht.
Ekzistojnë artiodaktil ditore , crepuskulare dhe nate . Modeli i zgjimit të disa specieve ndryshon sipas stinës ose habitatit.
Riprodhimi dhe cikli jetsor
RedaktoNë përgjithësi, dythundrakët kanë tendencë të kenë periudha të gjata të shtatzanis , madhësi më të vogël pjellash dhe të posalindur më shumë të zhvilluar. Ashtu si me shumë gjitarë të tjerë, speciet në rajone të butë ose polare kanë një sezon fikse çiftëzimi , ndërsa ato në zona tropikale rriten gjatë gjithë vitit. Ata kryejnë sjellje poliogjene çiftëzimi, do të thotë një mashkull me disa femra dhe shtyp gjithë konkurrencën.
Gjatësia e periudhës së shtatzanis ndryshon nga katër në pesë muaj për derrat, drerët dhe dreri e myshqeve; gjashtë deri në dhjetë muaj për hipopotamët, dre dhe bovidët; dhjetë deri trembëdhjetë muaj për devetë; dhe katërmbëdhjetë deri në pesëmbëdhjetë muaj për gjirafat. Shumica japin një ose dy foshnje, por disa derra mund të japin deri në dhjetë.
Të porsalindurit janë pararendës (lindin relativisht të pjekur) dhe vijnë me sy të hapur dhe janë me flokë (me përjashtim të hipopotamëve pa flokë). Drerët dhe derrat e të miturve kanë pallto me shirita ose me njolla; modeli zhduket ndërsa plaken. Të miturit e disa specieve i kalojnë javët e para me nënën e tyre në një vend të sigurt, ku të tjerët mund të vrapojnë dhe të ndjekin tufën brenda disa orësh ose ditësh.
Jetëgjatësia është zakonisht njëzet deri në tridhjetë vjet; si në shumë gjitarë, speciet më të vogla shpesh kanë një jetëgjatësi më të shkurtër se speciet më të mëdha. Artiodaktilët me jetëgjatësinë më të gjatë janë hipopotamët, lopët dhe devetë, të cilat mund të jetojnë 40 deri në 50 vjet.
Grabitqarët dhe parazitët
RedaktoArtiodaktilët kanë grabitqarë natyrorë të ndryshëm në varësi të madhësisë dhe habitatit të tyre. Ekzistojnë disa mishngrënës që gjuajnë kafshë të tilla, duke përfshirë macet e mëdha (p.sh., luanë ) dhe arinjtë . Grabitqarët e tjerë janë krokodilët , ujqërit , shpendët grabitqar e mëdhenj dhe për speciet e vogla dhe kafshët e reja, gjarpërinjtë e mëdhenj .
Parazitët përfshijnë nematodë , pleshtat , morrat , apo fluket , por ato kanë efekte dobësuese vetëm kur infektimi është i rëndë.
Mardhëniet me njerzit
RedaktoZbutja
Redakto
Artiodaktilët janë gjuajtur nga njerëzit primitivë për arsye të ndryshme: për mish ose lesh , si dhe për të përdorur kockat dhe dhëmbët e tyre si armë ose mjete. Zbutja e tyre filloi rreth vitit 8000 pes. Deri më sot, njerëzit kanë zbutur dhitë, delet, lopët, devet, lamat, alpakata dhe derrat . Fillimisht, bagëtia përdorej kryesisht për ushqim, por ato filluan të përdoren për aktivitete pune rreth 3000 pes pes.[36] Ekzistojnë prova të qarta të antilopës që përdoren për ushqim 2 milion vjet më parë në Grykën e Olduvai , pjesë e Luginës së Madhe Rift . Cro-Magnons mbështetej shumë te renë për ushqim, lëkurë, mjete dhe armë; me uljen e temperaturave dhe rritjen e numrave të renëve në fund të Pleistocenit, ata u bënë pre e zgjedhjes.[36][37] Renet llogariten për 94% të eshtrave dhe dhëmbëve të gjetura në një shpellë mbi lumin Céou që ishte e banuar rreth 12.500 vjet më parë.[38]
Sot, artiodaktilat mbahen kryesisht për mishin, qumështin dhe leshin . Lopët shtëpiake, buallicat , jakët dhe devetë përdoren për punë ose si kafshë udhëtimi .[39]
Kërcënimet
Redakto
Niveli i rrezikimit të secilës dythundrak është i ndryshëm. Disa specie janë sinantropike (siç është derri i egër ) dhe janë përhapur në zona për të cilat nuk janë indigjenë, ose janë sjellë si kafshë në fermë ose kanë ikur si kafshë shtëpiake të njerëzve. Disa artiodaktil gjithashtu përfitojnë nga fakti se grabitqarët e tyre (p.sh. tigri tasmanian ) ishin degjeneruar rëndë nga gjuetia, të cilët i shihnin si konkurrencë.[35]
Në të kundërt, shumë artiodaktil kanë rënë ndjeshëm në numër, dhe disa madje janë zhdukur, kryesisht për shkak të gjuetisë së tepërt , dhe, së fundmi, shkatërrimit të habitatit . Specie të zhdukur përfshijnë disa gazela (të tilla si gazela arabe ), hipopotami i malagasit , antilopa blu , dhe drerin e Shomburgut . Dy lloje, oriksi i Skimitarit dhe dreri i At Davidit , janë zhdukur në të egër . Katërmbëdhjetë specie konsiderohen të rrezikuara në mënyrë kritike , duke përfshirë adaksin , kouprein, deven e egër bakteriane ,gazelën e Przevalskit , saigën , dhe derrin xhuxh. Njëzet e katër specie konsiderohen të rrezikuara .[40][41]
Referime
Redakto- ^ a b c Montgelard, Claudine; Catzeflis, Francois M.; Douzery, Emmanuel (1997). "Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences" (PDF). Molecular Biology and Evolution. 14 (5): 550–559. doi:10.1093/oxfordjournals.molbev.a025792. PMID 9159933.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Jessica M Theodor; Jörg Erfurt; Grégoire Métais (2007-10-23). "The earliest artiodactyls: Diacodexeidae, Dichobunidae, Homacodontidae, Leptochoeridae and Raoellidae". përmbledhur nga Donald R. Prothero; Scott E. Foss (red.). Evolution of Artiodactyls. Johns Hopkins University. fq. 32–58. ISBN 9780801887352.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ a b c d Spaulding, M; O'Leary, MA; Gatesy, J (2009). Farke, Andrew Allen (red.). "Relationships of Cetacea (Artiodactyla) Among Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution". PLoS ONE. 4 (9): e7062. Bibcode:2009PLoSO...4.7062S. doi:10.1371/journal.pone.0007062. PMC 2740860. PMID 19774069.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!)Mirëmbajtja CS1: DOI i lirë i pashënjuar (lidhja) - ^ Groves, Colin P.; Grubb, Peter (2011). Ungulate Taxonomy. Baltimore, Maryland: Johns Hopkins University Press. fq. 25. ISBN 978-1-4214-0093-8.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Graur, Dan; Higgins, Desmond G. (1994). "Molecular Evidence for the Inclusion of Cetaceans within the Order Artiodactyla" (PDF). Molecular Biology and Evolution: 357–364. Arkivuar nga origjinali (PDF) më 5 mars 2016. Marrë më 23 gusht 2015.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Gatesy, John; Hayashi, Cheryl; Cronin, Mathew A.; Arctander, Peter (1996). "Evidence from milk casein genes that cetaceans are close relatives of hippopotamid artiodactyls". Molecular Biology and Evolution. 13 (7): 954–963. doi:10.1093/oxfordjournals.molbev.a025663. PMID 8752004.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Shimamura, M. (1997). "Molecular evidence from retroposons that whales form a clade within even-toed ungulates". Nature. 388 (6643): 666–670. doi:10.1038/41759. PMID 9262399.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!)
- ^ Gatesy, John (1997). "More DNA Support for a Cetacea/Hippopotamidae Clade: The Blood-Clotting Protein Gene y-Fibrinogen". Molecular Biology and Evolution. 14 (5): 537–543. doi:10.1093/oxfordjournals.molbev.a025790. PMID 9159931.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ a b Agnarsson, Ingi; May-Collado, Laura J. (2008). "The phylogeny of Cetartiodactyla: The importance of dense taxon sampling, missing data, and the remarkable promise of cytochrome b to provide reliable species-level phylogenies". Molecular Phylogenetics and Evolution (në anglisht). 48 (3): 964–85. doi:10.1016/j.ympev.2008.05.046. PMID 18590827.
- ^ Gatesy, John; Milinkovitch, Michel; Waddell, Victor; Stanhope, Michael (1999). "Stability of Cladistic Relationships between Cetacea and Higher-Level Artiodactyl Taxa". Systematic Biology (në anglisht). 48 (1): 6–20. doi:10.1080/106351599260409. PMID 12078645.
- ^ Madsen, Ole; Willemsen, Diederik; Ursing, Björn M.; Arnason, Ulfur; de Jong, Wilfried W. (2002). "Molecular Evolution of the Mammalian Alpha 2B Adrenergic Receptor". Molecular Biology and Evolution. 19 (12): 2150–2160. doi:10.1093/oxfordjournals.molbev.a004040. PMID 12446807.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Amrine-Madsen, Heather; Koepfli, Klaus-Peter; Wayne, Robert K.; Springer, Mark S. (2003). "A new phylogenetic marker, apolipoprotein B, provides compelling evidence for eutherian relationships". Molecular Phylogenetics and Evolution (në anglisht). 28 (2): 225–240. doi:10.1016/s1055-7903(03)00118-0. PMID 12878460.
- ^ Savage, R. J. G.; Long, M. R. (1986). Mammal Evolution: an illustrated guide. New York: Facts on File. fq. 208. ISBN 978-0-8160-1194-0.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ a b Price, Samantha A.; Bininda-Emonds, Olaf R. P.; Gittleman, John L. (2005). "A complete phylogeny of the whales, dolphins and even-toed hoofed mammals (Cetartiodactyla)". Biological Reviews (në anglisht). 80 (3): 445–73. doi:10.1017/s1464793105006743. PMID 16094808.
- ^ etwa noch bei Nowak (1999) oder Hendrichs (2004)
- ^ Malcolm C. McKenna; Susan K. Bell (1997). 'Classification of Mammals - Above the Species Level. Columbia University Press. ISBN 978-0-231-11013-6.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Beck, N.R. (2006). "A higher-level MRP supertree of placental mammals". BMC Evol Biol. 6: 93. doi:10.1186/1471-2148-6-93. PMC 1654192. PMID 17101039.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!)Mirëmbajtja CS1: DOI i lirë i pashënjuar (lidhja) - ^ O'Leary, M.A.; Bloch, J.I.; Flynn, J.J.; Gaudin, T.J.; Giallombardo, A.; Giannini, N.P.; Goldberg, S.L.; Kraatz, B.P.; Luo, Z.-X.; Meng, J.; Ni, X.; Novacek, M.J.; Perini, F.A.; Randall, Z.S.; Rougier, G.W.; Sargis, E.J.; Silcox, M.T.; Simmons, N.B.; Spaulding, M.; Velazco, P.M.; Weksler, M.; Wible, J.R.; Cirranello, A.L. (2013). "The Placental Mammal Ancestor and the Post-K-Pg Radiation of Placentals". Science. 339 (6120): 662–667. doi:10.1126/science.1229237.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Song, S.; Liu, L.; Edwards, S.V.; Wu, S. (2012). "Resolving conflict in eutherian mammal phylogeny using phylogenomics and the multispecies coalescent model". Proceedings of the National Academy of Sciences. 109 (37): 14942–14947. doi:10.1073/pnas.1211733109.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ dos Reis, M.; Inoue, J.; Hasegawa, M.; Asher, R.J.; Donoghue, P.C.J.; Yang, Z. (2012). "Phylogenomic datasets provide both precision and accuracy in estimating the timescale of placental mammal phylogeny". Proceedings of the Royal Society B: Biological Sciences. 279 (1742): 3491–3500. doi:10.1098/rspb.2012.0683.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Upham, N.S.; Esselstyn, J.A.; Jetz, W. (2019). "Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation". PLOS Biology. 17 (12): e3000494. doi:10.1371/journal.pbio.3000494.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!)(see e.g. Fig S10) - ^ Wilson, D. E.; Reeder, D. M., red. (2005). Mammal Species of the World (bot. 3rd). Johns Hopkins University Press. fq. 111–184. ISBN 978-0-8018-8221-0.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Cui, P.; Ji, R.; Ding, F.; Qi, D.; Gao, H.; Meng, H.; Yu, J.; Hu, S.; Zhang, H. (2007). "A complete mitochondrial genome sequence of the wild two-humped camel (Camelus bactrianus ferus): an evolutionary history of Camelidae". BMC Genomics. 8 (1): 241. doi:10.1186/1471-2164-8-241. PMC 1939714. PMID 17640355.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!)Mirëmbajtja CS1: DOI i lirë i pashënjuar (lidhja) - ^ "Claws Out: Things You Didn't Know About Claws". Thomson Safaris (në anglishte amerikane). 2014-01-07. Marrë më 2016-09-24.
- ^ Franz-Viktor Salomon (2008). musculoskeletal system. In: F.-V. Salomon and others (eds.): Anatomy for veterinary medicine. fq. 22–234. ISBN 978-3-8304-1075-1.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ DeMiguel, Daniel; Azanza, Beatriz; Morales, Jorge (2014). "Key innovations in ruminant evolution: a paleontological perspective". Integrative Zoology. 9 (4): 412–433. doi:10.1111/1749-4877.12080. PMID 24148672.
{{cite journal}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Janis, C.M.; Scott, K.M. (1987). "The Interrelationships of Higher Ruminant Families with Special Emphasis on the Members of the Cervoidea". American Museum Novitates (në anglisht). 2893: 1–85. Arkivuar nga origjinali më 6 tetor 2014. Marrë më 4 shtator 2020.
- ^ a b Janis, C.; Jarman, P. (1984). Macdonald, D. (red.). The Encyclopedia of Mammals. New York: Facts on File. fq. 498–499. ISBN 978-0-87196-871-5.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Shively, C. L.; etj. (1985). "Some Aspects of the Nutritional Biology of the Collared Peccary". The Journal of Wildlife Management (në anglisht). 49 (3): 729–732. doi:10.2307/3801702. JSTOR 3801702.
- ^ Hewitt, David G (2011-06-24). Biology and Management of White-tailed Deer. ISBN 9781482295986.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Spinage, C. A. "Reproduction in the Uganda defassa waterbuck, Kobus defassa ugandae Neumann." Journal of reproduction and fertility 18.3 (1969): 445-457.
- ^ Yong, Hwan-Yul. "Reproductive System of Giraffe (Giraffa camelopardalis). Arkivuar 25 prill 2019 tek Wayback Machine" Journal of Embryo Transfer 24.4 (2009): 293-295.
- ^ Sumar, Julio. "Reproductive physiology in South American Camelids." Genetics of Reproduction in Sheep (2013): 81.
- ^ Uwe Gille (2008). urinary and sexual apparatus, urogenital Apparatus. In: F.-V. Salomon and others (eds.): Anatomy for veterinary medicine. fq. 368–403. ISBN 978-3-8304-1075-1.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ a b Pough, F. W.; Janis, C. M.; Heiser, J. B. (2005) [1979]. "Major Lineages of Mammals". Vertebrate Life (bot. 7th). Pearson. fq. 539. ISBN 978-0-13-127836-3.
{{cite book}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ a b "Artiodactyl". Encyclopædia Britannica Online (në anglisht). Encyclopædia Britannica, Inc. 2008. Marrë më 2008-10-17.
- ^ McKie, Robin (2012-09-22). "Humans hunted for meat 2 million years ago". the Guardian (në anglisht). Marrë më 2015-10-26.
- ^ "Bones From French Cave Show Neanderthals, Cro-Magnon Hunted Same Prey". ScienceDaily. 2003. Marrë më 17 tetor 2008.
{{cite web}}: Mungon ose është bosh parametri|language=(Ndihmë!) - ^ Clay, J. (2004). World Agriculture and the Environment: A Commodity-by-Commodity Guide to Impacts and Practices (në anglisht). Washington, D.C., USA: Island Press. ISBN 978-1-55963-370-3.
- ^ "Cetartiodactyla" (në anglisht). Marrë më 12 mars 2007.
- ^ "Artiodactyla" (në anglisht). Encyclopedia of Life. Marrë më 15 nëntor 2014.