
Diplodocus
Pour la grue ferroviaire, voir Diplodocus (chemin de fer).
Ne doit pas être confondu avec Diplocaulus.

Espèces de rang inférieur
- † Diplodocus carnegii Hatcher[2], 1901, nouvelle espèce type proposée en 2016[3]
- † Diplodocus hallorum D. Gillette[4], 1991
- † Diplodocus longus Marsh[1], 1878, nomen dubium, espèce type originale
Diplodocus est un genre fossile de très grands dinosaures herbivores sauropodes de la famille des diplodocidés ayant vécu au Jurassique supérieur (Kimméridgien supérieur), il y a environ entre 154 et 152 Ma (millions d'années), en Amérique du Nord où il a été découvert dans les parties moyenne et supérieure de la formation de Morrison dans les États de l'ouest des États-Unis.
Les premiers fossiles ont été découverts en 1877 par Samuel Wendell Williston dans les sédiments alluviaux ou marins peu profonds de la formation de Morrison. Diplodocus est l'un des fossiles de dinosaures les plus communs de cette formation ; il vivait à la même époque que d'autres sauropodes gigantesques comme Camarasaurus, Barosaurus, Apatosaurus et Brachiosaurus[5].
Diplodocus figure parmi les dinosaures les plus facilement identifiables ; c'est le plus connu des sauropodes, avec son long cou et sa longue queue en forme de fouet, ses quatre pattes robustes et sa tête plate. Sa grande taille peut avoir été un système de dissuasion contre les attaques de prédateurs comme Allosaurus et Ceratosaurus, dont les restes ont été trouvés dans la même strate, ce qui indique qu'ils ont coexisté avec Diplodocus.
Étymologie
[modifier | modifier le code]Le nom générique, inventé par Othniel Charles Marsh en 1878, est un néologisme latin dérivé du grec διπλόος (diploos) « double » et δοκός (dokos) « faisceau »[6], en raison de la présence d'os appelés arcs hémaux ou chevrons ayant une structure en double poutre et qui forment une longue succession osseuse dans la partie inférieure de la queue. On croyait initialement que ces os étaient caractéristiques des Diplodocus, mais ils ont été découverts depuis chez d'autres genres de la famille et chez des sauropodes non diplodocidés comme Mamenchisaurus.
Description
[modifier | modifier le code]
Diplodocus hallorum est en violet.

C'est un très grand quadrupède au long cou, avec une longue queue en forme de fouet. Ses membres antérieurs sont légèrement plus courts que ses membres postérieurs, ce qui lui donnait une posture horizontale. Le long cou, la longue queue et les quatre pattes robustes le font, mécaniquement, ressembler à un pont suspendu[7].
Taille
[modifier | modifier le code]L'espèce Diplodocus carnegii est décrite à partir d'un squelette complet[7]. Sa longueur totale est évaluée à 25 mètres par Gregory Paul en 2010[8] ; selon lui, l'estimation de sa masse varie entre 10 et 16 tonnes[8].
La seconde espèce valide, Diplodocus hallorum, connue à partir de restes fossiles partiels, était plus grande, mais l'estimation de sa taille est plus délicate. Sa longueur totale a été historiquement surévaluée avec une valeur de 54 mètres extrapolée à partir de restes partiels de Seismosaurus hallorum, ancien nom de D. hallorum, par l'inventeur de cette espèce, David Gillette en 1991[4]. Cette longueur en faisait alors le plus long dinosaure connu à l'exclusion du genre Amphicoelias, évalué toutefois à partir de restes encore plus fragmentaires .
Thomas Holtz en 2011 estime la taille de D. hallorum à 30 mètres, après que Kenneth Carpenter a constaté en 2006 que D. Gillette avait positionné trop en arrière certaines vertèbres caudales et que la treizième vertèbre de la queue de D. carnegii, qui servait de modèle pour évaluer la longueur de D. hallorum, appartenait en fait à un autre spécimen[9]. La masse de D. hallorum pouvait atteindre 24 tonnes[10].
Squelette
[modifier | modifier le code]Crâne
[modifier | modifier le code]
Aucun crâne complet clairement attribuable à Diplodocus n'a été identifié, bien qu'il soit envisagé comme plat et allongé. La tête de Diplodocus est très petite par rapport à la taille de l'animal, ce qui est une caractéristique chez les sauropodes. Diplodocus avait de petites dents pointues en forme de pinces orientées vers l'avant, uniquement présentes dans la partie antérieure de la gueule[12].
Narines
[modifier | modifier le code]
a) crâne montrant les ouvertures nasales au sommet du crâne,
b) dessin classique de la tête avec les narines sur le dessus de la tête,
c) avec une trompe hypothétique,
d) représentation moderne avec des narines à l'extrémité du museau et une possible chambre de résonance.
La tête du Diplodocus a été historiquement représentée avec les narines très hautes sur sa tête en raison de la position des ouvertures nasales au sommet du crâne. Robert T. Baker s'est demandé en 1986 si Diplodocus avait une trompe[14]. Une étude de 2006, conduite par P. Knoll et ses colègues a montré qu'il n'y avait aucune preuve paléoneuroanatomique pour une telle existence[15]. Il faut noter que le nerf facial est gros chez un animal avec une trompe, comme l'éléphant, car il doit innerver toute la trompe. Or les données paléontologiques suggèrent que le nerf facial était de très petite taille chez Diplodocus. Des études menées par Lawrence Witmer (2001) indiquent que, alors que les ouvertures nasales sont haut situées sur la tête, les narines étaient situés beaucoup plus bas sur le museau et qu'entre les deux pouvait éventuellement se trouver une chambre de résonance[16].
Cou
[modifier | modifier le code]
Son cou était composé d'au moins quinze vertèbres. Sa longueur pouvait dépasser 6 mètres[17], ce qui constitue un avantage compétitif indéniable pour l'accès à plus de nourriture, soit en hauteur, soit en projection horizontale, en particulier pour des plantes aquatiques de marais comme des massifs de prêles[18].
La posture du cou, à savoir s'il pouvait se redresser jusqu'à la verticale ou, au contraire, s'il ne pouvait pas se lever plus haut que l'horizontale, est un sujet qui fait toujours débat parmi les paléontologues. Une étude de Matthew J. Cobley et de ses collègues en 2013 pour les diplodocidés, conclut que leur flexibilité ne peut pas être basée uniquement sur l'ostéologie. En l'absence d'informations sur les tissus musculaires, les résultats sont à considérer avec prudence, même s'ils notent que les cous de sauropodes étaient probablement moins flexibles qu'on ne le pensait auparavant[19].
Mike P. Taylor, en 2014, remet en cause ces résultats après une étude des cous de Diplodocus et d'Apatosaurus. Il considère que les cartilages inter-vertébraux assuraient la flexibilité de leurs cous qui pouvaient se redresser au-delà de la verticale, même si la posture naturelle basée sur la morphologie des vertèbres, la pose neutre ostéologique, était horizontale avec la tête relevée en position d'alerte[20].
Chez les sauropodes, des postures du cou extrêmement différentes ont été envisagées, d'un cou incliné vers l'avant en position pour brouter uniquement la végétation au sol[21], jusqu'à des cous pouvant atteindre et même dépasser la verticale pour accéder aux hautes frondaisons comme chez les genres Brachiosaurus, Euhelopus et Kaatedocus[22],[23].
Pour Diplodocus, même dans l'hypothèse d'un cou peu flexible et horizontal, l'accès aux feuilles hautes de la couronne des arbres lui était toutefois permis en se redressant sur ses pattes arrière et en s'appuyant sur sa queue[18].
Queue
[modifier | modifier le code]

New Mexico Museum of Natural History and Science.
Diplodocus a une queue extrêmement longue, composée d'environ 80 vertèbres caudales[24], soit presque le double du nombre de vertèbres de queue de certains sauropodes comme Shunosaurus (43) ou Camarasaurus (53). La queue servait vraisemblablement de contrepoids au cou.
Le dessous de la partie antérieure de la queue est caractérisé par la présence d'os appelés arcs hémaux ou chevrons qui forment, dans la partie centrale de la queue, un double faisceau d'excroissances qui a donné son nom au genre. Ils peuvent avoir fourni un appui pour les vertèbres, ou peut-être protéger les vaisseaux sanguins de l'écrasement lorsque la lourde queue de l'animal était posée sur le sol. Ces os sont également rencontrés chez certains dinosaures apparentés[25].
La partie postérieure de la queue est dépourvue de ces os ; elle devient très fine et prend une forme de fouet. Différentes hypothèses ont été avancées sur le rôle de cette queue, qu'il soit défensif ou utilisé lors de combats intraspécifiques ou de parades amoureuses[26]. Diplodocus aurait fait claquer sa queue comme le fouet d'un cocher pour se défendre ou seulement pour produire un bruit sec et caractéristique de reconnaissance ou de parade. En 1997, il a été calculé qu'un tel fouet pouvait provoquer un bang supersonique de plus de 200 décibels[27].
La découverte en 1990 d'empreintes de peau de diplodocidés a montré que certaines espèces portaient des épines kératineuses étroites et pointues comme celles des iguanes. Ces épines peuvent atteindre 18 centimètres de long, elles sont placées sur la partie arrière (fouet) de la queue et, peut-être, sur l'ensemble du dos et du cou de l'animal[28],[29]. Ces empreintes de peau, découvertes sur le site de Howe Quarry près du village de Shell dans le Wyoming, ont été trouvées à proximité de restes fossiles de diplodocidés ressemblant à Dipolodocus et Barosaurus[28]. Cependant les spécimens découverts sur ce site ont depuis été révisés et attribués à Kaatedocus siberi et à Barosaurus sp., plutôt qu'à Diplodocus[23],[11].
Membres
[modifier | modifier le code]Comme chez les autres sauropodes, l'équivalent de la main était fortement modifié, les os des doigts et la paume étant disposés en une colonne verticale, en fer à cheval en coupe transversale. Diplodocus n'avait pas de griffes à l’extrémité des cinq doigts mais des coussinets, sauf au niveau d'un doigt qui portait une griffe exceptionnellement longue par rapport à celles des autres sauropodes, aplatie latéralement, et séparée des os de la main. La fonction de cette griffe particulièrement spécialisée reste inconnue.
Tissus mous
[modifier | modifier le code]La découverte dans le Montana, de fragments de peau fossilisés et attribués au genre Diplodocus, suggère que ceux ci arboraient des écailles et nodosités de différentes tailles selon l'emplacement de celles-ci sur le corps[30].
Les différentes espèces
[modifier | modifier le code]Historique
[modifier | modifier le code]Après la première découverte par Benjamin Mudge et Samuel Wendell Williston en 1877 à Cañon City dans le Colorado du squelette nommé Diplodocus longus par Othniel Charles Marsh en 1878[1], plusieurs espèces de Diplodocus ont été décrites entre 1878 et 1924.
D. carnegii, décrite en 1901, est l'espèce la mieux connue, par de nombreux squelettes post-crâniens, bien qu'elle ne soit pas l'espèce type et qu'aucun crâne de cette espèce n'ait été clairement été rattaché au reste du squelette. L'espèce est surtout très célèbre par le grand nombre de moulages existant dans des musées du monde entier[25].
D. hayi, découverte en 1902 dans le Wyoming par William H. Utterback est connue par un crâne et un squelette post-crânien partiel. Elle est décrite en 1924 par W. J. Holland[31]. En 2015, elle est renommée et attribuée à un nouveau genre, Galeamopus, comme c'est le cas pour beaucoup de spécimens de « Diplodocus », ce qui fait qu'aucun crâne n'est attribué avec certitude à Diplodocus[11],[13].
Deux genres de sauropodes de la formation de Morrison, Diplodocus et Barosaurus, ont des os des membres postérieurs très similaires. Dans le passé, de nombreux os isolés de membres ont été automatiquement attribués à Diplodocus, mais peuvent, en fait, avoir appartenu à Barosaurus[32].
Espèces valides
[modifier | modifier le code]

Seules deux espèces de Diplodocus sont considérées comme valides, D. carnegii et D. hallorum. Le statut de l'espèce type originelle D. longus est incertain, considéré comme un nomen dubium.
Diplodocus carnegii (également orthographié D. carnegiei), du nom de l'industriel et philanthrope américain Andrew Carnegie, est la mieux connue, principalement en raison de la découverte d'un squelette quasi complet collecté par Jacob Wortman, du Carnegie Museum of Natural History à Pittsburgh, en Pennsylvanie et décrit et nommé par John Bell Hatcher en 1901. En 2016, Emanuel Tschopp et Octavio Mateus ont déposé une requête auprès de la commission internationale de nomenclature zoologique pour que D. carnegii soit retenue comme nouvelle espèce type du genre Diplodocus en remplacement de D. longus, dont les fossiles sont considérés par la plupart des paléontologues comme des nomen dubia, ou appartenant à une autre espèce[11],[3].
Diplodocus hallorum, a été découverte en 1979 sous la forme d'un squelette partiel (référencé NMMNH P-3690) comprenant des vertèbres, des côtes et un pelvis. Elle a été décrite en 1991 par D. Gillette sous le nom de Seismosaurus halli[4]. George Olshevsky a par la suite proposé de la renommer S. hallorum, invoquant une erreur grammaticale de l'inventeur, ce dont D. Gillette s'est acquitté en 1994[33] et qui a ensuite été repris par la plupart des paléontologues dont K. Carpenter[9].
En 2004, lors d'une présentation à l'assemblée annuelle de la société américaine de géologie, Spencer Lucas et ses collègues défendent l'idée que Seismosaurus devrait être considéré comme un synonyme junior du genre Diplodocus[34]. Ceci a été suivi par une publication plus détaillée en 2006, qui non seulement rebaptisait l'espèce Diplodocus hallorum, mais émettait aussi l'hypothèse qu'il pourrait s'agir de la même espèce que D. longus[35]. La supposition que D. hallorum doive être considéré comme un spécimen de D. longus a également été reprise par les auteurs d'une redescription de Supersaurus, réfutant une hypothèse précédente qui considérait Seismosaurus et Supersaurus comme étant les mêmes genres[36].
En 2015, l'analyse phylogénétique des diplodocidés conduite par E. Tschopp et ses collègues conclut que les spécimens les plus complets de D. longus étaient bien de la même espèce que D. hallorum, et que les autres attribués à D. longus devaient être considérés comme des nomen dubia[11].
Espèces réattribuées ou douteuses
[modifier | modifier le code]- D. longus, l'espèce type, est connue principalement par deux crânes et une série de vertèbres caudales trouvés dans la formation de Morrison dans le Colorado et l'Utah[17]. Ces fossiles sont aujourd'hui attribués à D. hallorum ou des nomen dubia[11].
- D. lacustris a été nommée par O. C. Marsh en 1884, dans le contexte singulier de la guerre des os avec E. D. Cope, à partir des restes d'un animal de petite taille de la formation de Morrison, dans le Colorado[37]. Ces restes ont été ensuite considérés comme étant ceux d'un animal immature, plutôt que ceux d'une espèce distincte. L'espèce était classée comme un nomen dubium[17]. En 2015 cependant, E. Tschopp et ses collègues réassignent ce spécimen au genre Camarasaurus[11].
Paléobiologie
[modifier | modifier le code]Grâce à une multitude de restes de squelettes, Diplodocus est l'un des dinosaures les mieux connus. De nombreux aspects de son mode de vie ont été les sujets des diverses théories au fil des ans.
Habitat
[modifier | modifier le code]Jusqu'au milieu du XXe siècle, il était supposé que Diplodocus vivait dans l'eau, en raison de la position de ses narines au sommet du crâne[37],[2]. On a souvent représenté les autres grands sauropodes tels que Brachiosaurus et Apatosaurus ayant un mode de vie similaire.
En 1951, une étude de Kenneth A. Kermack montre que ces sauropodes n'auraient probablement pas pu respirer par les narines lorsque le reste du corps aurait été immergé, car la pression de l'eau sur la paroi thoracique aurait été trop forte[38]. Depuis les années 1970, un consensus général a fait des sauropodes des animaux bien terrestres, broutant les arbres, les fougères et les buissons.
Posture
[modifier | modifier le code]
La représentation de la posture de Diplodocus a considérablement changé au fil du temps. Par exemple, un dessin de reconstruction classique de 1910 par Oliver P. Hay représente deux Diplodocus avec les membres écartés sur les côtés de leurs corps comme des lézards sur les bords d'une rivière. Hay prétendait que Diplodocus marchait un peu comme un lézard avec les jambes largement écartées[40],[39] et sa conception était soutenue par le célèbre paléontologue allemand Gustav Tornier. Cependant, cette hypothèse a été contestée dès 1910 par W.J. Holland qui a démontré qu'un Diplodocus marchant ainsi aurait eu besoin d'une tranchée pour faire avancer son ventre[41]. Des empreintes de sauropodes trouvées dans les années 1930 ont fini par réfuter la théorie de Hay.
Alimentation
[modifier | modifier le code]Diplodocus avait une denture très inhabituelle comparée à celle des autres sauropodes. Les couronnes dentaires sont longues et fines, elliptiques en coupe transversale, tandis que le sommet forme une pointe émoussée de forme triangulaire[12]. La facette d'usure la plus importante est sur le sommet contrairement aux zones d'usure observées chez d'autres sauropodes où elles sont situées sur la face labiale (joue) à la fois pour les dents du haut et du bas[12].
Les diplodocidés avaient donc un mécanisme d'alimentation radicalement différent de celui des autres sauropodes. Il semble qu'il mordait les branches des arbres et que leurs dents servaient de râteau ou de pinces, et qu'il arrachait les feuilles en tirant la tête en arrière[42],[43],[44], ce qui expliquerait les traces d'usures inhabituelles (résultant du contact des dents avec les aliments). Pour effeuiller les branches, une rangée de dents aurait servi à séparer les feuilles de la tige, tandis que l'autre aurait servi de guide et de stabilisateur. Upchurch en 2000 souligne que la région préorbitaire (en avant des yeux) allongée de son crâne, devait lui permettre d'effeuiller en une seule action de grandes longueurs de branches[12]. En outre un mouvement vers l'arrière de la mâchoire inférieure (« mouvement palinal ») pourrait avoir joué deux rôles importants dans son comportement alimentaire : 1) une augmentation de l'ouverture de la gueule et 2) un ajustement précis de la position relative des deux rangées des dents, permettant un meilleur effeuillage[12]. Comme pour la plupart des sauropodes, des gastrolithes ont été retrouvés à proximité des os de Diplodocus. Ces pierres avalées facilitaient le broyage des fibres végétales les plus résistantes et ainsi aidaient à la digestion.
En 2012, Mark Young et ses collègues réalisent des modélisations biomécaniques du crâne des diplodocinés. Ils en concluent que 1) l'hypothèse que les dents étaient utilisées pour arracher l'écorce (wikt:écorçage) des arbres n'est pas possible car cela aurait soumis les dents et le crâne à des contraintes trop fortes, 2) par contre, les hypothèses d'effeuillage des branches par les dents et/ou d'ajustement précis des dents étaient plausibles d'un point de vue biomécanique[45].
Les dents des diplodocinés étaient continuellement remplacées au cours de leurs vies. Michael D'Emic et ses collègues en 2013 ont observé que dans chaque cavité dentaire, jusqu'à cinq dents de remplacement pouvaient se développer pour remplacer la dent active. Ils estiment que le remplacement s'effectuait habituellement en moins de 35 jours[46]. Des études sur les dents de diplodocinés révèlent également que ceux-ci préféraient une végétation différente de celle aux autres sauropodes découverts en leur compagnie dans la formation de Morrison, comme Camarasaurus. Cette adaptation à différents types de végétations aurait pu permettre aux diverses espèces de sauropodes de coexister sans compétition pour l'accès à la nourriture[46].
Diplodocus, avec son cou plus ou moins flexible latéralement et de haut en bas, mais aussi la possibilité d'utiliser sa queue comme point d'appui et de se redresser sur ses pattes arrière (appui tripode), avait la possibilité de manger à plusieurs niveaux de végétation, du sol jusqu'à environ 10 mètres de hauteur[47]. L’amplitude des mouvements du cou lui aurait aussi permis de brouter, conduisant certains scientifiques à spéculer sur le fait que Diplodocus pouvait pâturer des prairies flottantes ou immergées, à partir des berges. Ce concept de régime alimentaire est étayé par la longueur relative entre les membres antérieurs et postérieurs. En outre, ses dents en pointes pouvaient être utilisées pour consommer des plantes d'eau douce[48].
En 2010, Jeffrey Whitlock et ses collègues décrivent le crâne d'un Diplodocus juvénile (référencé CM 11255) qui diffère sensiblement des crânes adultes du même genre[49]. C'est le seul crâne connu d'un diplodociné juvénile[23]. Son museau est plus rond que chez les adultes, ses dents sont présentes plus à l'arrière des mâchoires et ses orbites sont un peu plus grandes[23]. Ces différences suggèrent que les adultes et les juvéniles se nourrissaient de manière différente. Une telle disparité écologique entre les adultes et les jeunes n'avait pas été observée auparavant chez les sauropodomorphes[49].
Reproduction et croissance
[modifier | modifier le code]À ce jour, aucun nid fossilisé attribué à Diplodocus n'a été retrouvé. En revanche, d'autres sauropodes comme Titanosaurus et Saltasaurus ont été associés à des sites de nidification fossilisés[50],[51]. Les sites de nidification de Titanosaurus indiquent que ces animaux devaient pondre leurs œufs en groupe sur une grande surface dans de nombreux trous peu profonds creusés dans le sol, chacun d'entre eux étant couvert par la végétation. Il est possible que Diplodocus ait eu le même comportement. Le documentaire « Sur la terre des dinosaures » dépeint une femelle Diplodocus pondant ses œufs à l'aide d'une tarière (ovipositeur), mais cela n'est que pure spéculation de la part du réalisateur.
Après un certain nombre d'études d'histologie osseuse, on sait que Diplodocus, ainsi que les autres sauropodes, grandissait à un rythme très rapide, atteignant sa maturité sexuelle en un peu plus d'une décennie, tout en continuant de grandir tout au long de sa vie[52],[53],[54]. On pensait auparavant que les sauropodes continuaient de croître lentement tout au long de leur vie, leur arrivée à maturité sexuelle prenant des décennies.
Pour Diplodocus et d'autres sauropodes, la taille des couvées et des œufs individuels étaient étonnamment petites pour de tels animaux. Cela semble avoir été une adaptation aux pressions exercées par la prédation, étant donné que des œufs de grande taille auraient nécessité un temps d'incubation plus long augmentant ainsi le risque de prédation[55].
Classification
[modifier | modifier le code]Diplodocus est le genre type de la famille des Diplodocidae[37]. Les membres de cette famille, tout en étant des animaux assez massifs, étaient nettement plus élancés que les autres sauropodes, tels que les titanosauridés et les brachiosauridés. Ils se caractérisent par un long cou et une longue queue tenus en position horizontale, avec des membres antérieurs plus courts que les membres postérieurs. Les diplodocidés ont prospéré au Jurassique supérieur, en Amérique du Nord, où ils sont connus par de très nombreux spécimens et un grand nombre d'espèces, et en Afrique[24]. À partir des années 2010, de nouveaux genres de diplodocidés ont été décrits ou réattribués en Europe (Supersaurus)[11], en Amérique du Nord (Kaatedocus et Galeamopus) et en Amérique du Sud (Leinkupal), ce dernier étant le premier diplodocidé découvert à avoir vécu après le Jurassique, au Crétacé[56].
Une sous-famille, les Diplodocinae, regroupe le genre Diplodocus et ses plus proches parents, Barosaurus, Galeamopus, Leinkupal, Supersaurus, Tornieria et Kaatedocus[11],[13].
Un peu plus éloignés mais contemporains, les célèbres genres Apatosaurus et Brontosaurus sont quant à eux placés dans l'autre sous-famille des diplodocidés, les Apatosaurinae[11].
Le diplodocidés sont rattachés au clade des Diplodocoidea avec deux autres familles, les dicraeosauridés, les rebbachisauridés[57],[11]. Ce clade est le groupe frère des Macronaria au sein des Neosauropoda, le plus grand, le plus diversifié et le plus riche clade des sauropodomorphes.
Cladogramme
[modifier | modifier le code]Le cladogramme ci-dessous, établi en 2017 par Emmanuel Tschopp et Octavio Mateus[13] et modifiant légèrement celui de Tschopp et ses collègues de 2015[11], montre la position des deux espèces de Diplodocus comme les plus évoluées des diplodocinés :
| Diplodocidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Diplodocus dans les musées
[modifier | modifier le code]



Diplodocus est un dinosaure célèbre et très souvent représenté car c'est le dinosaure sauropode le plus exposé dans des lieux publics. Ceci est probablement dû en grande partie à l'abondance de ses restes fossiles et à son ancien statut de plus long des dinosaures. Le don de moulages de squelettes montés par l'industriel et philanthrope Andrew Carnegie à de nombreux musées du monde entier au début du XXe siècle a beaucoup fait pour familiariser l'animal[14],[58].
Des moulages de squelettes de Diplodocus sont toujours exposés dans de nombreux musées à travers le monde, y compris un exceptionnel D. hayi au musée des sciences naturelles de Houston.
Les musées qui possèdent un squelette de D. carnegii sont :
- Le Musée d'histoire naturelle Carnegie à Pittsburgh possède le squelette original, monté en 1904 et inauguré en 1907 ;
- le Musée d'histoire naturelle de Londres, première réplique, dévoilée le 12 mai 1905 ;
- le Musée d'histoire naturelle de Berlin, en Allemagne, réplique dévoilée au début de mai 1908 ;
- le Muséum national d'histoire naturelle à Paris, en France, réplique dévoilée le 15 juin 1908 ;
- le Muséum de Vienne, en Autriche, réplique dévoilée en 1909 ;
- le Musée de paléontologie et de géologie de Bologne, en Italie, réplique dévoilée en 1909. Des copies du crâne de cette réplique (c'est-à-dire de « deuxième génération »), sont exposées dans les musées de Milan et Naples ;
- le Musée de zoologie de l'Académie russe des sciences à Saint-Pétersbourg, en Russie, réplique dévoilée en 1910 ;
- le Musée de la Plata, à La Plata près de Buenos Aires, en Argentine, réplique dévoilée en 1912 ;
- le Musée national des sciences naturelles d'Espagne, à Madrid, réplique dévoilée au début de 1914 ;
- le Musée de Paléontologie de Mexico, réplique dévoilée en 1929 ;
- le Musée de paléontologie de Munich, en Allemagne, réplique offerte en 1932 mais jamais montée.
Un autre D. carnegii est exposé au Musée d'histoire naturelle de Chicago. En outre, un D. longus est exposé au Musée Senckenberg à Francfort (squelette composé de plusieurs spécimens, donnés en 1907 par l'American Museum of Natural History), en Allemagne. Un squelette monté et plus complet de D. longus se trouve au Musée national d'histoire naturelle de Washington, tandis qu'un squelette monté de D. hallorum (anciennement Seismosaurus), qui est peut-être un D. longus peut être vu au Musée d'histoire naturelle et des sciences du Nouveau-Mexique.
Diplodocus dans la culture populaire
[modifier | modifier le code]Diplodocus a été un thème fréquent dans les films de dinosaures, à la fois documentaires et de fiction, à commencer par Gertie le dinosaure de Winsor McCay en 1914. Il a été présenté dans le deuxième épisode de la série télévisée Sur la trace des Dinosaures. L'épisode Le temps des Titans retrace la vie d'un diplodocus il y a 152 millions d'années. Le livre de James A. Michener, Colorado saga, a un chapitre consacré à un diplodocus, racontant la vie et la mort d'un individu.
Diplodocus est une figure souvent trouvée dans les dinosaures en jouets et en maquette. Il y a eu deux éditions picturales du diplodocus de la collection Carnegie.
La fiancée du diplodocus est une planche (No 662) de l'imagerie Pellerin d'Épinal. Elle raconte une farce cruelle jouée par deux loustics à une brave provinciale venue à Paris consulter une agence matrimoniale.
Dans la bande dessinée Le Sceptre d'Ottokar de la série Les Aventures de Tintin, créée par le dessinateur belge Hergé, Milou dérobe un os du squelette de l'espèce fictive Diplodocus gigantibus exposé dans le musée d'histoire naturelle de Klow[59].
Diplodocus est le nom communément donné, probablement en raison de son aspect général, à l'EPTVF (Engin Poseur de travées de Voie Ferrée), anciennement exploité par le 5e Régiment du Génie, aujourd'hui classé en France.
Galerie d'images
[modifier | modifier le code]-

Dessin de D. carnegii par Nobu Tamura -
![Moulage d'un crâne d'un diplodociné indéterminé (CM 11161)[11], attribué précédemment à D. longus au Smithsonian museum of Natural History, Washington DC](//upload.wikimedia.org/wikipedia/commons/thumb/c/c1/Diplodocus_skull.jpg/234px-Diplodocus_skull.jpg) Moulage d'un crâne d'un diplodociné indéterminé (CM 11161)[11], attribué précédemment à D. longus au Smithsonian museum of Natural History, Washington DC
Moulage d'un crâne d'un diplodociné indéterminé (CM 11161)[11], attribué précédemment à D. longus au Smithsonian museum of Natural History, Washington DC -

Os chevron de la partie antérieure de la queue. Os dépourvu des deux excroissances qui n’existent que dans la partie médiane de la queue -

Squelette de la tête et du cou de Diplodocus, Smithsonian museum of Natural History, Washington D.C. -

Ancienne représentation, sans doute erronée -

Diplodocus
(Charles R. Knight, 1912) -
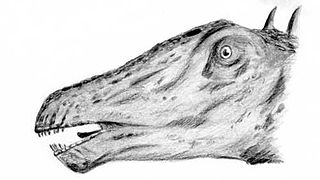
Tête -

Crâne

![Moulage d'un crâne d'un diplodociné indéterminé (CM 11161)[11], attribué précédemment à D. longus au Smithsonian museum of Natural History, Washington DC](/https/fr.wikipedia.org/wiki/Fichier:Diplodocus_skull.jpg)





Voir aussi
[modifier | modifier le code]Références taxinomiques
[modifier | modifier le code]- (en) Référence Paleobiology Database : Diplodocus Marsh, 1878
- (en) Référence Paleobiology Database : Diplodocus carnegii Hatcher, 1901
- (en) Référence Paleobiology Database : Diplodocus hallorum Gillette, 1991
Annexes
[modifier | modifier le code]Articles connexes
[modifier | modifier le code]Notes et références
[modifier | modifier le code]- (en) O.C. Marsh, « Principal Characters of American Jurassic Dinosaurs », American Journal of Science, vol. 16, no 95, , p. 412–414 (lire en ligne)
- (en) Hatcher JB. "Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton,". Memoirs of the Carnegie Museum, vol. 1 (1901), p. 1–63
- (en) E. Tschopp et O. Mateus, « Diplodocus Marsh, 1878 (Dinosauria, Sauropoda): proposed designation of D. carnegii Hatcher, 1901 as the type species », Bulletin of Zoological Nomenclature, vol. 73, no 1, , p. 17–24 (DOI 10.21805/bzn.v73i1.a22)
- (en) D.D. Gillette, « Seismosaurus halli, gen. et sp. nov., a new sauropod dinosaur from the Morrison Formation (Upper Jurassic/Lower Cretaceous) of New Mexico, USA », Journal of Vertebrate Paleontology, vol. 11, no 4, , p. 417–433 (DOI 10.1080/02724634.1991.10011413)
- (en) Christine C.E. & Peterson, F. (2004). "Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis". Sedimentary Geology 167, 309–355
- Skarlatos Vyzantios (Σκαρλάτος Βυζάντιος).Dictionnaire grec-français (1856)
- (en) Lambert D. (1993)The Ultimate Dinosaur Book (ISBN 0-86438-417-3). De fait, Diplodocus est le plus long dinosaure connu à partir d'un squelette complet
- (en) Gregory S. Paul, Princeton Field Guide to Dinosaurs, Princeton, Princeton University Press, , 320 p. (ISBN 978-0-691-13720-9)
- (en) Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131–138
- (en) Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- (en) Emanuel Tschopp, Octavio Mateus & Roger B.J. Benson (2015), « A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda) », PeerJ 3:e857; DOI 10.7717/peerj.857 https://peerj.com/articles/857/
- (en) Upchurch, P. et Barrett, P.M., Evolution of Herbivory in Terrestrial Vertebrates : perspectives from the fossil record, Cambridge, Cambridge University Press, , 256 p. (ISBN 0-521-59449-9), « The evolution of sauropod feeding mechanism »
- (en) Emanuel Tschopp et Octávio Mateus, « Osteology of Galeamopus pabsti sp. nov. (Sauropoda: Diplodocidae), with implications for neurocentral closure timing, and the cervico-dorsal transition in diplodocids. », PeerJ, vol. 5, , e3179 (DOI 10.7717/peerj.3179)
- (en) Robert T. Bakker, The Dinosaur Heresies : New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction, New York, William Morrow, , poche (ISBN 978-0-14-010055-6)
- F. Knoll, P.M. Galton, R. López-Antoñanzas, (2006). "Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus." Geobios, 39: 215–221
- (en) Lawrence M. Witmer et al., "Nostril Position in Dinosaurs and other Vertebrates and its Significance for Nasal Function." Science 293, 850 (2001)
- (en) Upchurch P, Barrett PM, Dodson P, The Dinosauria (2nd Edition), University of California Press, (ISBN 0-520-24209-2), « Sauropoda », p. 305
- (en) P. Martin Sander, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, Eva-Maria Griebeler, Hanns-Christian Gunga, Jürgen Hummel, Heinrich Mallison, Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas Tütken, Oliver Wings et Ulrich Witzel, « Biology of the sauropod dinosaurs: the evolution of gigantism », Biological Reviews, vol. 86, no 1, , p. 117–155 (ISSN 1464-7931, PMID 21251189, PMCID 3045712, DOI 10.1111/j.1469-185X.2010.00137.x)
- (en) M. J. Cobley, E. J. Rayfield et P. M. Barrett, « Inter-Vertebral Flexibility of the Ostrich Neck : Implications for Estimating Sauropod Neck Flexibility », PLoS ONE, vol. 8, , e72187 (PMID 23967284, DOI 10.1371/journal.pone.0072187)
- (en) M.P. Taylor, Quantifying the effect of intervertebral cartilage on the neutral posture in the necks of sauropod dinosaurs, vol. 2, , e712 (PMID 25551027, DOI 10.7717/peerj.712, lire en ligne)
- (en) Stevens, K.A. & Parrish, M. (1999). "Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs". Science 284, 798–800
- (en) A. Christian et G. Dzemski, Biology of the sauropod dinosaurs : Understanding the life of giants, Bloomington, Indiana University Press, , « Neck posture in sauropods »
- (en) E. Tschopp et O. V. Mateus, « The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs », Journal of Systematic Palaeontology, vol. 11, , p. 1 (DOI 10.1080/14772019.2012.746589)
- (en) JA Wilson, The Sauropods : Evolution and Paleobiology, Berkeley (Calif.) etc., Indiana University Press, , 349 p. (ISBN 0-520-24623-3), « Overview of Sauropod Phylogeny and Evolution », p. 15–49
- (en) Michael J. Benton, Prehistoric Life, Dorling Kindersley, , 512 p. (ISBN 978-0-7566-9910-9)
- (en) WJ Holland, « Heads and Tails: a few notes relating to the structure of sauropod dinosaurs. », Annals of the Carnegie Museum, vol. 9, , p. 273–278
- (en) Nathan P. Myhrvold, « Supersonic sauropods? Tail dynamics in the diplodocids », Paleobiology, vol. 23, no 4,
- (en) S. A. Czerkas, « Discovery of dermal spines reveals a new look for sauropod dinosaurs », Geology, vol. 20, , p. 1068–1070 (DOI 10.1130/0091-7613(1992)020<1068:dodsra>2.3.co;2)
- (en) Czerkas, S. A. (1994). "The history and interpretation of sauropod skin impressions." In Aspects of Sauropod Paleobiology (M. G. Lockley, V. F. dos Santos, C. A. Meyer, and A. P. Hunt, Eds.), Gaia No. 10. (Lisbon, Portugal)
- T. Gallagher, J. Poole, J. P. Schein, Evidence of integumentary scale diversity in the late Jurassic Sauropod Diplodocus sp. from the Mother’s Day Quarry, Montana, PeerJ, 29 Avril 2021
- (en) Holland WJ. The skull of Diplodocus. Memoirs of the Carnegie Museum IX; 379–403 (1924)
- (en) McIntosh, Thunder Lizards : The Sauropodomorph Dinosaurs, Indiana University Press, (ISBN 0-253-34542-1), « The Genus Barosaurus (Marsh) », p. 38–77
- (en) Gillette, D.D., 1994, Seismosaurus: The Earth Shaker. New York, Columbia University Press, 205 pp
- (en) Lucas S, Herne M, Heckert A, Hunt A, and Sullivan R. Reappraisal of Seismosaurus, A Late Jurassic Sauropod Dinosaur from New Mexico. The Geological Society of America, 2004 Denver Annual Meeting (November 7–10, 2004)
- (en) SG Lucas, JA Spielman, LA Rinehart et al., Paleontology and Geology of the Upper Morrison Formation, New Mexico Museum of Natural History and Science (bulletin 36), , « Taxonomic status of Seismosaurus hallorum, a Late Jurassic sauropod dinosaur from New Mexico », p. 149–161
- (en) D.M. Lovelace, S.A. Hartman et W.R. Wahl, « Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny », Arquivos do Museu Nacional, vol. 65, no 4, , p. 527–544 (ISSN 0365-4508, CiteSeerx 10.1.1.603.7472, lire en ligne)
- (en) O.C. Marsh, « Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidae, a new family of the Sauropoda », American Journal of Science, vol. 3, , p. 160–168
- (en) Kenneth A. Kermack, « A note on the habits of sauropods », Annals and Magazine of Natural History, vol. 12, no 4, , p. 830–832
- (en) Hay, O. P., 1910, Proceedings of the Washington Academy of Sciences, vol. 12, p. 1–25
- (en) Hay, Oliver P., "On the Habits and Pose of the Sauropod Dinosaurs, especially of Diplodocus." The American Naturalist, Vol. XLII, Oct. 1908
- (en) W. J. Holland, "A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus carnegii in the Carnegie Museum", The American Naturalist, 44:259–283. 1910
- (en) D.B. Norman (1985). "The illustrated Encyclopedia of Dinosaurs". London: Salamander Books Ltd
- (en) P. Dodson (1990). Sauropod paleoecology. in: "The Dinosauria" 1st Edition, (Eds. Weishampel, D.B., Dodson, P. & Osmólska, H.)
- (en) P.M. Barrett & P. Upchurch (1994). Feeding mechanisms of Diplodocus. Gaia 10, 195–204
- (en) Mark T. Young, Emily J. Rayfield, Casey M. Holliday, Lawrence M. Witmer, David J. Button, Paul Upchurch et Paul M. Barrett, « Cranial biomechanics of Diplodocus (Dinosauria, Sauropoda) : testing hypotheses of feeding behaviour in an extinct megaherbivore », Naturwissenschaften, vol. 99, , p. 637–643 (ISSN 1432-1904, PMID 22790834, DOI 10.1007/s00114-012-0944-y)
- (en) M. D. D’Emic, J. A. Whitlock, K. M. Smith, D. C. Fisher et J. A. Wilson, « Evolution of high tooth replacement rates in sauropod dinosaurs », PLoS ONE, vol. 8, no 7, , e69235 (PMID 23874921, PMCID 3714237, DOI 10.1371/journal.pone.0069235)
- (en) P.M. Barrett & P. Upchurch (2005). Sauropodomorph Diversity through Time, Paleoecological and Macroevolutionary Implications. in : "The Sauropods: Evolution and Paleobiology" (Eds. Curry, K. C.)
- (en) Stevens KA, Parrish JM, Thunder Lizards : The Sauropodomorph Dinosaurs, Indiana University Press, , 212–232 p. (ISBN 0-253-34542-1), « Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs »
- (en) John A. Whitlock, Jeffrey A. Wilson et Matthew C. Lamanna, « Description of a Nearly Complete Juvenile Skull of Diplodocus (Sauropoda: Diplodocoidea) from the Late Jurassic of North America », Journal of Vertebrate Paleontology, 2, vol. 30, , p. 442–457 (DOI 10.1080/02724631003617647)
- (en) Walking on Eggs: The Astonishing Discovery of Thousands of Dinosaur Eggs in the Badlands of Patagonia, by Luis Chiappe and Lowell Dingus. 19 June 2001, Scribner
- (en) Grellet-Tinner, Chiappe, & Coria, "Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentina)", Can. J. Earth Sci. 41(8): 949–960 (2004)
- (en) Sander, P. M. (2000). "Long bone histology of the Tendaguru sauropods: Implications for growth and biology". Paleobiology 26, 466–488
- (en) Sander, P. M., N. Klein, E. Buffetaut, G. Cuny, V. Suteethorn, and J. Le Loeuff (2004). "Adaptive radiation in sauropod dinosaurs: Bone histology indicates rapid evolution of giant body size through acceleration". Organisms, Diversity & Evolution 4, 165–173
- (en) P.M. Sander and N. Klein (2005). "Developmental plasticity in the life history of a prosauropod dinosaur". Science 310 1800–1802
- (en) Ruxton, Graeme D., Birchard, Geoffrey F. et Deeming, D Charles, « Incubation time as an important influence on egg production and distribution into clutches for sauropod dinosaurs », Paleobiology, vol. 40, no 3, , p. 323–330 (DOI 10.1666/13028)
- (en) Pablo A. Gallina, Sebastián Apesteguía, Alejandro Haluza et Juan I. Canale, « A Diplodocid Sauropod Survivor from the Early Cretaceous of South America », PLoS ONE, vol. 9, no 5, , e97128 (PMID 24828328, PMCID 4020797, DOI 10.1371/journal.pone.0097128)
- (en) Taylor, M.P. et Naish, D., « The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda) », PaleoBios, vol. 25, no 2, , p. 1–7 (ISSN 0031-0298)
- (en) Rea, Tom, Bone Wars: The Excavation and Celebrity of Andrew Carnegie's Dinosaur. Pittsburgh: Pittsburgh University Press, 2001
- Hergé, Le Sceptre d'Ottokar, Tournai, Casterman, coll. « Les Aventures de Tintin », (1re éd. 1947), 62 planches, 30 cm (ISBN 2-2030-0107-0), planche 34.