Diagrama esquemático del ciclo de vida haplo-diploide (con alternancia de generaciones multicelulares). Referencias: n: generación haploide (gametofítica), 2n: generación diploide (esporofítica), m!: mitosis, M!: meiosis, F!: fecundación círculo: primer estadio del ciclo de vida, unicelular cuadrado: estadios siguientes del ciclo de vida, multicelulares.
La alternancia de generaciones (también conocida como metagénesis o heterogénesis) es un tipo de ciclo biológico de vida propio de la reproducción sexual, presente desde su ancestro común en todos los eucariotas (en algunos perdida secundariamente) caracterizado por una alternancia de fasesnucleares; es decir, a partir de la meiosis se pasa a una fase haploide y por fusión de los gametos se pasa a la fase diploide.
La alternancia de generaciones ocurre en aquellas plantas y algas en Archaeplastida y Heterokontophyta que tienen distintas etapas haploides sexuales y diploides asexuales. En estos grupos, un gametofito haploide multicelular con n cromosomas se alterna con un esporofito diploide multicelular con 2n cromosomas, formado por n pares. Un esporofito maduro produce esporas haploides por meiosis, un proceso que reduce el número de cromosomas a la mitad, de 2n a n.
En una alternancia de fases gamética, el organismo es diploide, ya que solo la generación diploide se divide por mitosis hasta volverse multicelular, y la meiosis se produce justo antes de formarse los gametos. El "individuo adulto" es diploide. Son diploides, como por ejemplo, la mayoría de los animales.
En una alternancia de fases cigótica, tras la formación del cigoto se produce la meiosis sin pasar por una etapa multicelular, y es la fase haploide la que se divide por mitosis hasta volverse multicelular, con lo cual el organismo es haploide. El "individuo adulto" es haploide.
En los haplodiplontes hay una fase haploide multicelular y una fase diploide multicelular. Por eso hay dos generaciones alternadas de individuos, una diplonte y una haplonte (hay alternancia de generaciones). El cigoto diploide se divide por mitosis para formar la generación diploide multicelular. En el individuo adulto diploide se produce la meiosis para producir esporas a partir de las cuales se forma el individuo adulto haploide por mitosis. Esta fase da lugar a los gametos que se fusionarán en un nuevo cigoto, reiniciando el ciclo. Este ciclo de vida se da en plantas, al individuo haplonte multicelular se lo llama gametófito y al individuo diplonte multicelular se lo llama esporófito. A veces tanto el haplonte adulto como el diplonte adulto son similares entre sí, y solo se los diferencia en el análisis genético y al aparecer sus estructuras reproductivas (como en la lechuga de mar o Ulva), entonces se dice que el ciclo de vida es haplodiplonte con alternancia "homofásica" de generaciones, o generaciones isomorfas. A veces el haplonte adulto y el diplonte adulto son muy diferentes entre sí, como pasa en las plantas terrestres, entonces se dice que el ciclo de vida es haplodiplonte con alternancia "heterofásica" de generaciones, o generaciones heteromorfas. Cuando tanto el gametófito como el esporófito son observables a simple vista, como pasa en los musgos y en los helechos, se dice que la alternancia de generaciones es bien manifiesta. En las plantas con semilla, solo la fase diplonte multicelular (esporófito) es observable a simple vista, el gametófito femenino (que da la gameta femenina) está encerrado dentro del óvulo (que luego se convierte en semilla), el gametófito masculino (que da la gameta masculina) está encerrado dentro del grano de polen.
La alternancia de generaciones se define como la alternancia de formas multicelulares diploides y haploides en el ciclo de vida del organismo, independientemente de si estas formas son de vida libre.[1] En algunas especies, como el alga Ulva lactuca, las formas diploide y haploide son de hecho organismos independientes de vida libre, esencialmente idénticos en apariencia y, por lo tanto, se dice que son isomorfos. Los gametos haploides que nadan libremente forman un cigoto diploide que germina en un esporofito diploide multicelular. El esporofito produce esporas haploides que nadan libremente por meiosis que germinan en gametofitos haploides.[2]
Sin embargo, en algunos otros grupos, el esporofito o el gametofito están muy reducidos y son incapaces de vivir libremente. Por ejemplo, en todas las briófitas la generación del gametofito es dominante y el esporofito depende de ella. Por el contrario, en todas las plantas terrestres vasculares modernas los gametofitos están fuertemente reducidos, aunque la evidencia fósil indica que se derivaron de ancestros isomórficos.[3] En plantas con semillas, el gametofito femenino se desarrolla totalmente dentro del esporofito, que lo protege y nutre, y del esporofito embrionario que produce. Los granos de polen, que son los gametofitos masculinos, se reducen a solo unas pocas células (solo tres células en muchos casos). Aquí la noción de dos generaciones es menos obvia; como dicen Bateman y Dimichele, [s] porofito y gametofito funcionan efectivamente como un solo organismo.[4] El término alternativo alternancia de fases puede ser entonces más apropiado.[5]
Los debates sobre la alternancia de generaciones a principios del siglo XX pueden ser confusos porque coexisten varias formas de clasificar las "generaciones" (sexual frente a asexual, gametofito frente a esporofito, haploide frente a diploide, etc.).[6]
Inicialmente, Chamisso y Steenstrup describieron la sucesión de generaciones organizadas de manera diferente (sexuales y asexuales) en animales como "alternancia de generaciones", mientras estudiaban el desarrollo de animales tunicados, cnidarios y trematodos.[6] Este fenómeno también se conoce como heterogamia. Actualmente, el término alternancia de generaciones se asocia casi exclusivamente con los ciclos de vida de las plantas, específicamente con la alternancia de gametofitos haploides y esporofitos diploides.[6]
Wilhelm Hofmeister demostró la alternancia morfológica de generaciones en las plantas,[7] entre una generación portadora de esporas (esporofito) y una generación portadora de gametos (gametofito).[8][9] En ese momento, surgió un debate centrado en el origen de la generación asexual de las plantas terrestres (es decir, el esporofito) y se caracteriza convencionalmente como un conflicto entre las teorías de antitético ( Čelakovský , 1874) y homólogo ( Pringsheim , 1876) alternancia de generaciones.[6] Čelakovský acuñó las palabras esporofito y gametofito.[cita requerida]
Eduard Strasburger (1874) descubrió la alternancia entre fases nucleares diploides y haploides,[6] también llamada alternancia citológica de fases nucleares.[10] Aunque la mayoría de las veces coinciden, la alternancia morfológica y la alternancia de fases nucleares a veces son independientes entre sí, por ejemplo, en muchas algas rojas , la misma fase nuclear puede corresponder a dos generaciones morfológicas diversas.[10] En algunos helechos que perdieron la reproducción sexual, no hay cambio en la fase nuclear, pero se mantiene la alternancia de generaciones.[11]
El diagrama anterior muestra los elementos fundamentales de la alternancia de generaciones en las plantas. Las numerosas variaciones que se encuentran en los diferentes grupos de plantas se describen mediante el uso de estos conceptos más adelante en el artículo. Empezando por la derecha del diagrama, los procesos implicados son los siguientes:[12]
Dos gametos unicelulares haploides, cada uno con n cromosomas no apareados, se fusionan para formar un cigoto unicelular diploide, que ahora contiene n pares de cromosomas, es decir, 2 n cromosomas en total.
El cigoto diploide unicelular germina y se divide por el proceso normal (mitosis), que mantiene el número de cromosomas en 2n. El resultado es un organismo diploide multicelular, llamado esporófito (porque en su madurez produce esporas).
Cuando alcanza la madurez, el esporofito produce uno o más esporangios (singular: esporangio) que son los órganos que producen las células madre diploides de las esporas (esporocitos). Estos se dividen mediante un proceso especial (meiosis) que reduce el número de cromosomas a la mitad. Esto da lugar inicialmente a cuatro esporas unicelulares haploides, cada una de las cuales contiene "n" cromosomas no apareados.
La espora unicelular haploide germina y se divide por el proceso normal (mitosis), que mantiene el número de cromosomas en n. El resultado es un organismo multicelular haploide, llamado gametofito (porque produce gametos en la madurez).
Cuando alcanza la madurez, el gametofito produce uno o más gametangios (singular: gametangio) que son los órganos que producen gametos haploides. Al menos un tipo de gameto posee algún mecanismo para llegar a otro gameto con el fin de fusionarse con él.
La "alternancia de generaciones" en el ciclo vital se produce, pues, entre una generación diploide (2n) de esporofitos y una generación haploide (n) de gametofitos.
Gametofito del helecho Onoclea sensibilis (el talo plano de la parte inferior) con un descendiente esporofito que empieza a crecer a partir de él (la pequeña fronda de la parte superior).
La situación es muy diferente a la de los animales, donde el proceso fundamental es que un individuo diploide (2n)
produce gametos haploides (n) por meiosis. En los animales no se producen esporas (es decir, células haploides capaces de sufrir mitosis), por lo que no hay generación multicelular asexual. Algunos insectos tienen machos haploides que se desarrollan a partir de huevos no fecundados, pero las hembras son todas diploides.
Gametofito de Mnium hornum, un musgo.Esporofito de Lomaria discolor, un helecho.
El diagrama mostrado arriba es una buena representación del ciclo vital de algunas algas multicelulares (por ejemplo, el género Cladophora) que tienen esporofitos y gametofitos de apariencia casi idéntica y que no tienen diferentes tipos de esporas o gametos.[13]
Sin embargo, hay muchas variaciones posibles en los elementos fundamentales de un ciclo de vida que tiene alternancia de generaciones. Cada variación puede ocurrir por separado o en combinación, lo que resulta en una desconcertante variedad de ciclos de vida. Los términos utilizados por los botánicos para describir estos ciclos vitales pueden ser igualmente desconcertantes. Como dicen Bateman y Dimichele "[...] la alternancia de generaciones se ha convertido en un marasmo terminológico; a menudo, un término representa varios conceptos o un concepto está representado por varios términos."[14]
Las posibles variantes son:
Importancia relativa del esporofito y del gametofito.
Igual (homomorfía or isomorfía). Las algas filamentosas del género Cladophora , que se encuentran predominantemente en agua dulce, tienen esporofitos diploides y gametofitos haploides que son indistinguibles externamente. [19] Ninguna planta terrestre viva tiene esporofitos y gametofitos igualmente dominantes, aunque algunas teorías de la evolución de la alternancia de generaciones sugieren que las plantas terrestres ancestrales sí lo hicieron.
Desigual (heteromorfía or anisomorfía).
Gametofito dominante (gametofito). En hepáticas, musgos y antocerotes, la forma dominante es el gametofito haploide. El esporofito diploide no es capaz de una existencia independiente, obtiene la mayor parte de su nutrición del gametofito padre y no tiene clorofila cuando madura.[15]
Esporofito dominante (esporofito). En los helechos, tanto el esporofito como el gametofito son capaces de vivir de forma independiente, pero la forma dominante es el esporofito diploide. El gametofito haploide es mucho más pequeño y de estructura más simple. En las plantas con semillas, el gametofito se reduce aún más (como mínimo a solo tres células), obteniendo toda su nutrición del esporofito. La reducción extrema en el tamaño del gametofito y su retención dentro del esporofito significa que cuando se aplica a las plantas con semillas, el término 'alternancia de generaciones' es algo engañoso: "[s]porofito y gametofito funcionan efectivamente como un solo organismo"[4] Some authors have preferred the term 'alternation of phases'.[5]
Diferenciación de los gametos.
Ambos gametos iguales (isogamia). Al igual que otras especies de Cladophora , C. callicoma tiene gametos flagelados que son idénticos en apariencia y capacidad de movimiento.
Gametos de dos tamaños distintos (anisogamia).
Ambos de similar motilidad .. Las especies de Ulva , la lechuga de mar, tienen gametos que tienen dos flagelos y, por lo tanto, son móviles. Sin embargo, son de dos tamaños: gametos 'femeninos' más grandes y gametos 'masculinos' más pequeños.[16]
Uno grande y sésil, otro pequeño y móvil (oogamia). Los megagametos más grandes y sésiles son los óvulos (huevos), y los microgametos más pequeños y móviles son los espermatozoides (espermatozoides). El grado de movilidad de los espermatozoides puede ser muy limitado (como en el caso de las plantas con flores), pero todos son capaces de moverse hacia los óvulos sésiles. Cuando (como ocurre casi siempre) los espermatozoides y los óvulos se producen en diferentes tipos de gametangios, los que producen espermatozoides se denominan anteridios (anteridio singular) y los que producen óvulos arquegonios (singular archegonium).Gametofito de Pellia epiphylla con esporofitos que crecen a partir de restos de archegonia..
Antheridia y archegonia ocurren en el mismo gametofito, que luego se llamamonoico. (Muchas fuentes, incluidas las relacionadas con los briófitos, usan el término "monoico" para esta situación y "dioico" para lo contrario.[17][18] Aquí, "monoico" y "dioico" se usan solo para los esporofitos). Pellia epiphylla tiene al gametofito como generación dominante. Es monoico: los pequeños anteridios rojizos que producen esperma se encuentran dispersos a lo largo de la nervadura central, mientras que los arquegonios que producen huevos crecen más cerca de las puntas de las divisiones de la planta.[19]
Antheridia y archegonia ocurren en diferentes gametofitos, que luego se denominandioicos.
El musgo Mnium hornum tiene el gametofito como generación dominante. Es dioico: las plantas masculinas producen solo anteridios en rosetas terminales, las plantas femeninas producen solo arquegonios en forma de cápsulas pedunculadas.[20] Los gametofitos de plantas con semillas también son dioicos. Sin embargo, el esporofito original puede ser monoico, produciendo gametofitos tanto masculinos como femeninos, o dioico, produciendo gametofitos de un solo género. Los gametofitos de las plantas con semillas tienen un tamaño extremadamente reducido; el arquegonio consta solo de un pequeño número de células, y el gametofito masculino completo puede estar representado por solo dos células.[21]
Ciclos de vida de los diferentes grupos de plantas
La alternancia de generaciones se produce en casi todas las algas multicelulares rojas y verdes, tanto en las formas de agua dulce (como la Cladophora) como en las algas marinas (como la Ulva). En la mayoría, las generaciones son homomórficas (isomórficas) y de vida libre. Algunas especies de algas rojas presentan una compleja alternancia trifásica de generaciones, en la que hay una fase gametofítica y dos fases esporofíticas distintas. Para más información, véase Algas rojas: Reproducción.
Todas las plantas terrestres tienen una alternancia heteromórfica (anisomórfica) de generaciones, en la que el esporofito y el gametofito son claramente diferentes. Todas las briofitas, es decir, las liverworts, los musgos y las hornworts, tienen la generación del gametofito como la más conspicua. Como ejemplo, consideremos un musgo monoico. En la planta madura (el gametofito) se desarrollan anteridios y arquegonios. En presencia de agua, los espermatozoides biflagelados de los anteridios nadan hasta la arquegonia y se produce la fecundación, lo que da lugar a un esporofito diploide. El esporofito crece a partir del arquegonio. Su cuerpo se compone de un largo tallo rematado por una cápsula en cuyo interior las células productoras de esporas sufren la meiosis para formar esporas haploides. La mayoría de los musgos dependen del viento para dispersar estas esporas, aunque Splachnum sphaericum es entomófilo, reclutando insectos para dispersar sus esporas. Para más información, véase Liverwort: Ciclo de vida, Moss: Ciclo de vida, Hornwort: Ciclo de vida.
Diagrama de alternancia de generaciones en hepáticas.
Diagrama del ciclo de vida del musgo
Diagrama del ciclo de vida del Hornwort
En los helechos y sus aliados, incluidos los musgos y las colas de caballo, la planta más llamativa que se observa en el campo es el esporofito diploide. Las esporas haploides se desarrollan en sori en el envés de las frondas y son dispersadas por el viento (o en algunos casos, flotando en el agua). Si las condiciones son adecuadas, una espora germina y crece hasta formar un cuerpo vegetal poco visible llamado prótalo. El prótalo haploide no se asemeja al esporófito, por lo que los helechos y sus aliados tienen una alternancia heteromórfica de generaciones. El prótalo es de corta duración, pero lleva a cabo la reproducción sexual, produciendo el cigoto diploide que luego crece fuera del prótalo como el esporofito. Para más información, véase Fern: Life cycle.
Diagrama de alternancia de generaciones en helechos.
Un gametofito (prótalo) de Dicksonia sp.
Un esporofito de Dicksonia antarctica.
Parte inferior de una fronda de Dicksonia antarctica que muestra los soros, o estructuras productoras de esporas.
En las espermatofitass, las plantas con semilla, el esporofito es la fase multicelular dominante; los gametofitos tienen un tamaño muy reducido y una morfología muy diferente. Toda la generación de gametofitos, con la única excepción de los granos de polen (microgametofitos), está contenida en el esporofito. El ciclo vital de una planta con flor dioica (angiosperma), el sauce, se ha descrito con cierto detalle en una sección anterior (Un ciclo vital complejo). El ciclo vital de una gimnosperma es similar. Sin embargo, las plantas con flor tienen además un fenómeno llamado 'doble fecundación'. Dos núcleos de espermatozoides de un grano de polen (el microgametofito), en lugar de un solo espermatozoide, entran en el arquegonio del megagametofito; uno se fusiona con el núcleo del óvulo para formar el cigoto, el otro se fusiona con otros dos núcleos del gametofito para formar el 'endospermo', que nutre al embrión en desarrollo.
↑Kerp, H.; Trewin, N.H.; Hass, H. (2003), «New gametophytes from the Lower Devonian Rhynie Chert», Transactions of the Royal Society of Edinburgh: Earth Sciences94 (4): 411-428, S2CID128629425, doi:10.1017/S026359330000078X.
↑ abFeldmann, J. (1972), «Les problèmes actuels de l'alternance de génerations chez les Algues», Bulletin de la Société Botanique de France(en francés)119: 7-38, doi:10.1080/00378941.1972.10839073.
Bell, P.R.; Hemsley, A.R. (2000), Green Plants: their Origin and Diversity (2nd edición), Cambridge, etc.: Cambridge University Press, ISBN978-0-521-64109-8.
Guiry, M.D.; Guiry, G.M. (2008), «Cladophora», AlgaeBase (World-wide electronic publication, National University of Ireland, Galway), consultado el 21 de julio de 2011.
Kirby, A. (2001), Ulva, the sea lettuce, Monterey Bay Aquarium Research Institute, archivado desde el original el 16 de mayo de 2011, consultado el 1 de enero de 2011.
Shyam, R. (1980), «On the life-cycle, cytology and taxonomy of Cladophora callicoma from India», American Journal of Botany67 (5): 619-24, JSTOR2442655, doi:10.2307/2442655.
Sporne, K.R. (1974a), The Morphology of Angiosperms, London: Hutchinson, ISBN978-0-09-120611-6.
Sporne, K.R. (1974b), The Morphology of Gymnosperms (2nd edición), London: Hutchinson, ISBN978-0-09-077152-3.
Stewart, W.N.; Rothwell, G.W. (1993), Paleobotany and the Evolution of Plants (2nd edición), Cambridge, UK: Cambridge University Press, ISBN978-0-521-38294-6.
Watson, E.V. (1981), British Mosses and Liverworts (3rd edición), Cambridge, UK: Cambridge University Press, ISBN978-0-521-28536-0.
Taylor, T.N.; Kerp, H.; Hass, H. (2005), «Life history biology of early land plants: Deciphering the gametophyte phase», Proceedings of the National Academy of Sciences of the United States of America102 (16): 5892-5897, PMC556298, PMID15809414, doi:10.1073/pnas.0501985102.







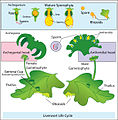 Diagrama de alternancia de generaciones en hepáticas.
Diagrama de alternancia de generaciones en hepáticas. Diagrama del ciclo de vida del musgo
Diagrama del ciclo de vida del musgo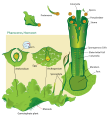 Diagrama del ciclo de vida del Hornwort
Diagrama del ciclo de vida del Hornwort










 Datos: Q725951
Datos: Q725951