Córtex motor primário
O córtex motor primário (área 4 de Brodmann) é uma região cerebral que, nos seres humanos, está localizada na porção dorsal do lobo frontal. É a região primária do sistema motor e trabalha em associação com outras áreas motoras, incluindo o córtex pré-motor, a área motora suplementar, o córtex parietal posterior e várias regiões do cérebro subcortical, para planejar e executar movimentos. O córtex motor primário é definido anatomicamente como a região do córtex que contém grandes neurônios conhecidos como células Betz. As células de Betz, juntamente com outros neurônios corticais, enviam axônios longos pela medula espinhal para sinapse nos circuitos interneurônios da medula espinhal e também diretamente nos neurônios motores alfa na medula espinhal que se conectam aos músculos.
| Córtex motor primário | |
|---|---|
 Área de Brodmann 4. | |
 Córtex motor primário marcado como córtex somatomotor. | |
| Detalhes | |
| Vascularização | artéria cerebral média; artéria cerebral anterior |
| Drenagem venosa | seio sagital superior |
| Identificadores | |
| Latim | cortex motorius primus |
| Gray | pág.821 |
| MeSH | Motor+Cortex |
No córtex motor primário, a representação do motor é organizada ordenadamente (de forma invertida) do dedo do pé (na parte superior do hemisfério cerebral) até a boca (na parte inferior) ao longo de uma dobra no córtex chamado sulco central. No entanto, algumas partes do corpo podem ser controladas por regiões parcialmente sobrepostas de córtex. Cada hemisfério cerebral do córtex motor primário contém apenas uma representação motora do lado oposto (contralateral) do corpo. A quantidade de córtex motor primário dedicado a uma parte do corpo não é proporcional ao tamanho absoluto da superfície do corpo, mas, em vez disso, à densidade relativa dos receptores motores cutâneos na referida parte do corpo. A densidade dos receptores motores cutâneos na parte do corpo é geralmente indicativa do grau necessário de precisão de movimento requerido na parte do corpo. Por esta razão, as mãos e o rosto humanos têm uma representação muito maior do que as pernas.
Para a descoberta do córtex motor primário e sua relação com outras áreas corticais do motor, veja o artigo principal sobre o córtex motor.
Estrutura
editar
O córtex motor primário humano está localizado na parede anterior do sulco central. Ele também se prolonga para fora do sulco parcialmente para o giro pré-centrado. Anteriormente, o córtex motor primário é limitado por um conjunto de áreas que se encontram no giro pré-central e que geralmente são consideradas para compor o córtex pré-motor lateral. Posteriormente, o córtex motor primário é delimitado pelo córtex somatossensorial primário, que fica na parede posterior do sulco central. Ventralmente, o córtex motor primário é limitado pelo córtex insular no sulco lateral. O córtex motor primário se estende dorsalmente até o topo do hemisfério e depois continua na parede medial do hemisfério.
A localização do córtex motor primário é mais óbvia no exame histológico devido à presença das células Betz distintivas. A camada V do córtex motor primário contém neurônios piramidais gigantes (70-100 μm) que são as células Betz. Esses neurônios enviam axônios longos aos núcleos motores contralaterais dos nervos cranianos e aos neurônios motores inferiores no chifre ventral da medula espinhal. Esses axônios fazem parte do trato corticoespinhal. As células de Betz representam apenas uma pequena porcentagem do trato corticoespinhal. Por algumas medidas, eles representam cerca de 10% dos neurônios do córtex motor primário que se projetam para a medula espinhal[1] ou cerca de 2-3% da projeção cortical total para a medula espinhal.[2] Embora as células de Betz não compõem a saída do motor inteiro do córtex, eles, no entanto, fornecem um marcador claro para o córtex motor primário. Esta região do córtex, caracterizada pela presença de células de Betz, foi denominada área 4 por Brodmann.
Caminho
editarÀ medida que os axônios motores viajam para baixo através da matéria branca cerebral, eles se aproximam e fazem parte do membro posterior da cápsula interna.
Eles continuam para baixo no tronco cerebral, onde alguns deles, depois de atravessar o lado contralateral, distribuem os núcleos motores do nervo craniano.
Depois de atravessar o lado contralateral na medula oblongada (decussação piramidal), os axônios viajam pela espinha medial como o trato corticoespinhal lateral.
As fibras que não atravessam o tronco encefálico viajam pelo trato corticoespinhal ventral separado, e a maioria deles passa para o lado contralateral na medula espinhal, pouco antes de atingir os neurônios motores inferiores.
Fornecimento de sangue
editarOs ramos da artéria cerebral média fornecem a maior parte do suprimento de sangue arterial para o córtex motor primário.
O aspecto medial (áreas das pernas) é fornecido por ramos da artéria cerebral anterior.
Função
editarHomúnculo
editarExiste uma representação amplamente somatotópica das diferentes partes do corpo no córtex motor primário em um arranjo chamado homúnculo motor (latim: pessoa pequena).[3] A área da perna está localizada perto da linha média, nas seções interiores da área do motor dobrando a fissura longitudinal medial. O lado lateral e convexo do córtex do motor primário é organizado de cima para baixo em áreas que correspondem às nádegas, torso, ombro, cotovelo, pulso, dedos, polegar, pálpebras, lábios e mandíbula. A área do braço e do motor de mão é a maior, e ocupa a parte do giro pré-central entre a área da perna e do rosto.
Essas áreas não são proporcionais ao seu tamanho no corpo com os lábios, as partes do rosto e as mãos representadas por áreas particularmente grandes. Após amputação ou paralisia, as áreas motoras podem mudar para adotar novas partes do corpo.
Entrada neural do tálamo
editarO córtex motor primário recebe entradas talâmicas de diferentes núcleos talâmicos. Entre outros:
- Núcleo lateral ventral para aferentes cerebelares
- Núcleo anterior ventral para aferentes de gânglios basais
Mapas alternativos
editar
Pelo menos duas modificações à ordem somatotópica clássica das partes do corpo foram relatadas no córtex motor primário de primatas.
Primeiro, a representação do braço pode ser organizada de forma central. No córtex dos macacos, os dígitos da mão estão representados em uma área central na borda posterior do córtex motor primário. Esta área central é cercada em três lados (nos lados dorsal, anterior e ventral) por uma representação das partes mais proximais do braço, incluindo o cotovelo e o ombro.[4][5] Nos seres humanos, a representação do dígito é cercada dorsalmente, anterior e ventralmente, por uma representação do pulso.[6]
Uma segunda modificação da ordem somatotópica clássica das partes do corpo é uma dupla representação dos dígitos e pulso estudados principalmente no córtex motor humano. Uma representação encontra-se numa região posterior chamada área 4p, e a outra encontra-se numa região anterior chamada área 4a. A área posterior pode ser ativada por atenção sem qualquer feedback sensorial e tem sido sugerida como importante para o início dos movimentos, enquanto a área anterior depende do feedback sensorial.[7] Também pode ser ativado por movimentos imaginários[8] dos dedos e ouvir a fala sem fazer movimentos reais. Esta área de representação anterior foi sugerida para ser importante na execução de movimentos envolvendo interações sensoriomotoras complexas.[9] É possível que a área 4a em seres humanos corresponda a algumas partes do córtex pré-motor caudal como descrito no córtex de macaco.
Em 2009, foi relatado, que existem duas regiões distintas evolutivas, uma mais antiga na superfície externa, e uma nova encontrada na fenda. O mais antigo se conecta aos neurônios da coluna vertebral através de interneurônios na medula espinhal. O mais recente, encontrado apenas em macacos, se conecta diretamente aos neurônios motores da coluna vertebral.[10] As conexões diretas após o nascimento são dominantes nas conexões indiretas e são mais flexíveis nos circuitos que podem desenvolver, o que permite o aprendizado pós-natal de habilidades motoras finas complexas. "O surgimento da" nova "região M1 durante a evolução da linhagem de primatas é, portanto, provável que tenha sido importante para a destreza manual aprimorada da mão humana".[11]
Equívocos comuns
editarCertos equívocos sobre o córtex motor primário são comuns em revisões secundárias, livros didáticos e material popular. Três dos equívocos mais comuns estão listados aqui.
Mapa segregado do corpo
editarUm dos equívocos mais comuns sobre o córtex motor primário é que o mapa do corpo é perfeitamente segregado. No entanto, não é um mapa de músculos individualizados ou mesmo partes do corpo individualizadas. O mapa contém uma sobreposição considerável. Essa sobreposição aumenta em regiões mais anteriores do córtex motor primário. Um dos principais objetivos da história do trabalho no córtex motor foi determinar o quanto as diferentes partes do corpo estão sobrepostas ou segregadas no córtex motor. Os pesquisadores que abordaram esta questão descobriram que o mapa da mão, braço e ombro continha ampla sobreposição.[3][5][6][12][13][14][15][16] Estudos que mapeiam a conectividade funcional precisa dos neurônios corticais aos músculos mostram que mesmo um único neurônio no córtex motor primário pode influenciar a atividade de muitos músculos relacionados a muitas articulações.[12] Em experimentos com gatos e macacos, à medida que os animais aprendem movimentos complexos e coordenados, o mapa no córtex motor primário se torna mais sobreposto, evidentemente aprendendo a integrar o controle de muitos músculos.[17][18] Em macacos, quando a estimulação elétrica é aplicada ao córtex motor em uma escala de tempo de comportamento, ele evoca movimentos complexos, altamente integrados, como alcançar a mão em forma de agarrar ou levar a mão à boca e abrir a boca.[19][20] Este tipo de evidência sugere que o córtex motor primário, ao mesmo tempo que contém um mapa áspero do corpo, pode participar na integração de músculos de maneiras significativas ao invés de segregar o controle de grupos musculares individuais. Sugeriu-se que um princípio mais profundo de organização pode ser um mapa das correlações estatísticas no repertório comportamental, em vez de um mapa das partes do corpo.[20][21] Na medida em que o repertório do movimento quebra em parte nas ações de partes do corpo separadas, o mapa contém um arranjo de corpo áspero e sobreposto.
M1 e córtex motor primário
editarO termo "M1" e o termo "córtex motor primário" são freqüentemente usados indistintamente. No entanto, eles vêm de diferentes tradições históricas e se referem a diferentes divisões do córtex. Alguns cientistas sugeriram que o córtex motor poderia ser dividido em uma tira de motor primária que era mais posterior e uma tira pré-operatória lateral mais anterior. Os primeiros pesquisadores que originalmente propuseram essa visão incluíram Campbell,[22] Vogt e Vogt[23] Foerster[24] e Fulton.[25] Outros sugeriram que o córtex motor não poderia ser dividido dessa maneira. Em vez disso, nesta segunda vista, o chamado primeiro motor primário e as tiras pré-motoras laterais compõem uma única área cortical denominada M1. Uma segunda área do motor na parede medial do hemisfério foi denominada M2 ou a área do motor suplementar. Os defensores desta visão incluíram Penfield[3] e Woolsey.[26] Atualmente, a distinção entre o córtex motor primário e o córtex pré-motor lateral é geralmente aceita. No entanto, o termo M1 às vezes é usado por engano para se referir ao córtex motor primário. Estritamente falando, M1 refere-se ao mapa único que, de acordo com alguns pesquisadores anteriores, engloba tanto o motor primário como o córtex pré-motor lateral.
Células Betz como o caminho comum final
editarAs células de Betz, ou células piramidais gigantes no córtex motor primário, às vezes são confundidas com a única ou principal saída do córtex para a medula espinhal. Esse erro é antigo, datando pelo menos de Campbell em 1905.[22] No entanto, as células de Betz compõe apenas cerca de 2-3% dos neurônios que se projetam do córtex para a medula espinhal[2] e apenas cerca de 10% dos neurônios que projetam especificamente de o córtex motor primário da medula espinhal.[1] Uma gama de áreas corticais, incluindo o córtex pré-motor, a área motora suplementar e mesmo o córtex somatossensorial primário, projetam a medula espinhal. Mesmo quando as células de Betz estão danificadas, o córtex ainda pode se comunicar com estruturas motoras subcorticais e controlar o movimento. Se o córtex motor primário com suas células Betz estiver danificado, os resultados temporários de paralisia e outras áreas corticais podem evidentemente assumir algumas das funções perdidas.
Significado clínico
editarAs lesões do giro pré-central resultam em paralisia do lado contralateral do corpo (paralisia facial, monoparesia braço/perna, hemiparesia).
Codificação de movimento
editarEvarts[27] sugeriu que cada neurônio no córtex motor contribui para a força em um músculo. À medida que o neurônio se torna ativo, ele envia um sinal para a medula espinhal, o sinal é retransmitido para um neurônio motor, o neurônio motor envia um sinal para um músculo e o músculo se contrai. Quanto mais atividade no neurônio do córtex motor, mais força muscular.
Georgopoulos e colegas[28][29][30] sugeriram que a força muscular sozinha era uma descrição muito simples. Eles treinaram macacos para chegar em várias direções e monitoraram a atividade dos neurônios no córtex motor. Eles descobriram que cada neurônio no córtex motor estava maximamente ativo durante uma direção específica de alcance e respondeu menos bem às direções vizinhas de alcance. Nesta base, eles sugeriram que os neurônios no córtex motor, ao "votar" ou ao agrupamento de suas influências em um "código de população", poderiam especificar precisamente uma direção de alcance.
A proposta de que os neurônios do córtex motor codificam a direção de um alcance tornou-se controversa. Scott e Kalaska[31] mostraram que cada neurônio do córtex motor estava melhor correlacionado com os detalhes do movimento das articulações e da força muscular do que com a direção do alcance. Schwartz e colegas[32] mostraram que os neurônios do córtex motor estavam bem correlacionados com a velocidade da mão. Strick e colegas[33] descobriram que alguns neurônios no córtex motor eram ativos em associação com a força muscular e alguns com a direção espacial do movimento. Todorov[34] propôs que as muitas correlações diferentes sejam o resultado de um controlador muscular em que muitos parâmetros de movimento estão correlacionados com a força muscular.
O código pelo qual os neurônios no córtex motor primado controlam a medula espinhal e, portanto, o movimento, continua a ser debatido.
Um progresso específico na compreensão de como o córtex motor causa movimento também foi feito no modelo de roedor. O córtex motor de roedores, como o córtex motor de macaco, pode conter sub-regiões que enfatizam diferentes tipos comuns de ações.[35][36] Por exemplo, uma região parece enfatizar o controle rítmico de whisking.[35][37][38] Neurônios nesta região projetam para um núcleo subcortical específico em que um gerador de padrão coordena o ritmo cíclico das bigodes. Este núcleo então projeta os músculos que controlam as bigodes.
Imagens adicionais
editar-
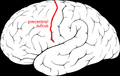 Sulco pré-central
Sulco pré-central -

-
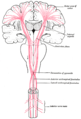 O trato do motor
O trato do motor



Veja também
editarReferências
- ↑ a b Rivara CB, Sherwood CC, Bouras C, Hof PR (2003). «Stereologic characterization and spatial distribution patterns of Betz cells in the human primary motor cortex». The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology. 270 (2): 137–151. PMID 12524689. doi:10.1002/ar.a.10015
- ↑ a b Lassek, A.M. (1941). «The pyramidal tract of the monkey». J. Comp. Neurol. 74: 193–202. doi:10.1002/cne.900740202
- ↑ a b c Penfield, W. and Boldrey, E. (1937). «Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation.». Brain. 60: 389–443. doi:10.1093/brain/60.4.389
- ↑ Kwan HC, MacKay WA, Murphy JT, Wong YC (1978). «Spatial organization of precentral cortex in awake primates. II. Motor outputs». J. Neurophysiol. 41: 1120–1131
- ↑ a b Park, M.C., Belhaj-Saif, A., Gordon, M. and Cheney, P.D. (2001). «Consistent features in the forelimb representation of primary motor cortex in rhesus macaques». J. Neurosci. 21: 2784–2792
- ↑ a b Meier, J.D., Aflalo, T.N., Kastner, S. and Graziano, M.S.A. (2008). «Complex organization of human primary motor cortex: A high-resolution fMRI study». J. Neurophysiol. 100: 1800–1812. PMC 2576195
 . PMID 18684903. doi:10.1152/jn.90531.2008
. PMID 18684903. doi:10.1152/jn.90531.2008
- ↑ Binkofski F, Fink GR, Geyer S, Buccino G, Gruber O, Shah NJ, Taylor JG, Seitz RJ, Zilles K, Freund HJ (2002). «Neural activity in human primary motor cortex areas 4a and 4b is modulated differentially by attention to action». J. Neurophysiol. 88 (1): 514–519. PMID 12091573
- ↑ Nikhil Sharma; P.S. Jones; T.A. Carpenter; Jean-Claude Baron (2008). «Mapping the involvement of BA 4a and 4p during Motor Imagery». NeuroImage. 41: 92–99. PMID 18358742. doi:10.1016/j.neuroimage.2008.02.009
- ↑ Terumitsu M, Ikeda K, Kwee IL, and Nakada, T (2009). «Participation of primary motor cortex area 4a in complex sensory processing: 3.0T fMRI study». NeuroReport. 20 (7): 679–683. PMID 19339906. doi:10.1097/WNR.0b013e32832a1820
- ↑ Rathelot, J.-A.; strick, P. L. (20 de janeiro de 2009). «Subdivisions of primary motor cortex based on cortico-motoneuronal cells». Proc. Natl. Acad. Sci. 106 (3): 918–923. PMC 2621250 . PMID 19139417. doi:10.1073/pnas.0808362106
- ↑ Costandi, Mo. «The evolution of manual dexterity». ScienceBlogs. Consultado em 29 de Novembro de 2015
- ↑ a b Cheney, P.D. & Fetz, E.E. (1985). «Comparable patterns of muscle facilitation evoked by individual corticomotoneuronal (CM) cells and by single intracortical microstimuli in primates: evidence for functional groups of CM cells». J. Neurophysiol. 53: 786–804
- ↑ Schieber, M.H. & Hibbard, L.S. (1993). «How somatotopic is the motor cortex hand area?». Science. 261 (5120): 489–492. PMID 8332915. doi:10.1126/science.8332915
- ↑ Rathelot, J.A. & Strick, P.L. (2006). «Muscle representation in the macaque motor cortex: an anatomical perspective». Proc. Natl. Acad. Sci. USA. 103: 8257–8262. PMC 1461407 . PMID 16702556. doi:10.1073/pnas.0602933103
- ↑ Sanes, J.N., Donoghue, J.P., Thangaraj, V., Edelman, R.R. and Warach, S. (1995). «Shared neural substrates controlling hand movements in human motor cortex». Science. 268 (5218): 1775–1777. PMID 7792606. doi:10.1126/science.7792606
- ↑ Donoghue, J.P., Leibovic, S. and Sanes, J.N. (1992). «Organization of the forelimb area in squirrel monkey motor cortex: representation of digit, wrist and elbow muscles». Exp. Brain Res. 89: 1–10. doi:10.1007/bf00228996
- ↑ Nudo, R.J., Milliken, G.W., Jenkins, W.M. and Merzenich, M.M. (1996). «Use-dependent alterations of movement representations in primary motor cortex of adult squirrel monkeys». J. Neurosci. 16: 785–807
- ↑ Martin, J.H., Engber, D. and Meng, Z. (2005). «Effect of forelimb use on postnatal development of the forelimb motor representation in primary motor cortex of the cat». J. Neurophysiol. 93 (5): 2822–2831. PMID 15574795. doi:10.1152/jn.01060.2004
- ↑ Graziano, M.S.A., Taylor, C.S.R. and Moore, T. (2002). «Complex movements evoked by microstimulation of precentral cortex». Neuron. 34 (5): 841–851. PMID 12062029. doi:10.1016/S0896-6273(02)00698-0
- ↑ a b Graziano, M.S.A. (2008). The Intelligent Movement Machine. Oxford, UK: Oxford University Press
- ↑ Graziano, M.S.A. and Aflalo, T.N. (2007). «Mapping behavioral repertoire onto the cortex.». Neuron. 56 (2): 239–251. PMID 17964243. doi:10.1016/j.neuron.2007.09.013
- ↑ a b Campbell, A. W. (1905). Histological Studies on the Localization of Cerebral Function. Cambridge, MA: Cambridge University Press
- ↑ Vogt, C. and Vogt, O. (1919). «Ergebnisse unserer Hirnforschung». Journal für Psychologie und Neurologie. 25: 277–462
- ↑ Foerster, O (1936). «The motor cortex of man in the light of Hughlings Jackson's doctrines». Brain. 59: 135–159. doi:10.1093/brain/59.2.135
- ↑ Fulton, J (1935). «A note on the definition of the "motor" and "premotor" areas». Brain. 58: 311–316. doi:10.1093/brain/58.2.311
- ↑ Woolsey, C.N., Settlage, P.H., Meyer, D.R., Sencer, W., Hamuy, T.P. and Travis, A.M. (1952). «Pattern of localization in precentral and "supplementary" motor areas and their relation to the concept of a premotor area». New York, NY: Raven Press. Association for Research in Nervous and Mental Disease. 30: 238–264
- ↑ Evarts, E.V. (1968). «Relation of pyramidal tract activity to force exerted during voluntary movement». J. Neurophysiol. 31 (1): 14–27. PMID 4966614
- ↑ Georgopoulos, A.P., Kalaska, J.F., Caminiti, R. and Massey, J.T. (1982). «On the relations between the direction of two-dimensional arm movements and cell discharge in primate motor cortex». J. Neurosci. 2 (11): 1527–1537. PMID 7143039
- ↑ Georgopoulos A.P., Kettner, R.E. and Schwartz, A.B. (1988). «Primate motor cortex and free arm movements to visual targets in three-dimensional space. II. Coding of the direction of movement by a neuronal population». J. Neurosci. 8 (8): 2928–2937. PMID 3411362
- ↑ Georgopoulos A.P., Schwartz, A.B. and Kettner, R.E. (1986). «Neuronal population coding of movement direction». Science. 233 (4771): 1416–1419. PMID 3749885. doi:10.1126/science.3749885
- ↑ Scott, S.H. & Kalaska, J.F. (1995). «Changes in motor cortex activity during reaching movements with similar hand paths but different arm postures». J. Neurophysiol. 73 (6): 2563–2567. PMID 7666162
- ↑ Moran, D.W. & Schwartz, A.B. (1999). «Motor cortical representation of speed and direction during reaching». J. Neurophysiol. 82 (5): 2676–2692. PMID 10561437
- ↑ Kakei, S., Hoffman, D. and Strick, P (1999). «Muscle and movement representations in the primary motor cortex». Science. 285 (5436): 2136–2139. PMID 10497133. doi:10.1126/science.285.5436.2136
- ↑ Todorov, E (2000). «Direct cortical control of muscle activation in voluntary arm movements: a model». Nat Neurosci. 3 (4): 391–398. PMID 10725930. doi:10.1038/73964
- ↑ a b Haiss, F. & Schwarz, C (2005). «Spatial segregation of different modes of movement control in the whisker representation of rat primary motor cortex». J. Neurosci. 25 (6): 1579–1587. PMID 15703412. doi:10.1523/JNEUROSCI.3760-04.2005
- ↑ Ramanathan, D., Conner, J.M. and Tuszynski, M.H. (2006). «A form of motor cortical plasticity that correlates with recovery of function after brain injury». Proc. Natl. Acad. Sci. U.S.A. 103 (30): 11370–11375. PMC 1544093 . PMID 16837575. doi:10.1073/pnas.0601065103
- ↑ Brecht, M., Schneider, M., Sakmann, B. and Margrie, T.W. (2004). «Whisker movements evoked by stimulation of single pyramidal cells in rat motor cortex». Nature. 427 (6976): 704–710. PMID 14973477. doi:10.1038/nature02266
- ↑ Cramer, N.P. & Keller, A (2006). «Cortical control of a whisking central pattern generator». J. Neurophysiol. 96 (1): 209–217. PMC 1764853 . PMID 16641387. doi:10.1152/jn.00071.2006