
Dinosourus
| Dinosourusse | |
|---|---|

| |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Subfilum: | |
| Klas: | |
| Subklas: | |
| Infraklas: | |
| Superorde: | Dinosauria (Owen, 1842)
|
| Belangrike groepe | |
| Dinosourusse | ||||||||
|---|---|---|---|---|---|---|---|---|
| Kladistiese klassifikasie | ||||||||
|
Dinosourusse of dinosouriërs was langer as 160 miljoen jaar die oorheersende landdiere, van laat in die Trias-periode (ongeveer 230 miljoen jaar gelede) tot aan die einde van die Kryt-periode (ongeveer 65 miljoen jaar gelede), toe die meeste uitgesterf het tydens die Kryt-Paleogeen-uitwissing. Fossielrekords dui daarop dat voëls tydens die Jura-periode uit Theropoda-dinosourusse ontwikkel het en die meeste paleontoloë beskou hulle as die enigste dinosourus-klade wat tot vandag nog lewe.[1]
Dinosourusse is ’n groep diere wat baie van mekaar verskil. Paleontoloë het met behulp van fossiele wêreldwyd reeds meer as 500 afsonderlike genera geïdentifiseer[2] wat saam meer as 1 000 spesies bevat.[3] Sommige dinosourusse was herbivore, terwyl ander karnivore was. Sommige was tweevoetig, ander viervoetig en nog ander kon tussen hierdie twee posture wissel. Baie spesies het uitvoerige skeletveranderings ondergaan, soos benerige pantsers, horings of kruine. Alhoewel hulle oor die algemeen as groot diere bekend is, was baie dinosourusse so groot soos mense of selfs kleiner. Die meeste het neste gebou en eiers gelê.
Die naam "dinosourus" is vir die eerste keer in 1842 deur sir Richard Owen gebruik en is afgelei van die Griekse deinos en sauros, letterlik "vreeslike" of "wonderlike akkedis". In die eerste helfte van die 20ste eeu het wetenskaplikes geglo dinosourusse was stadige, dom, koudbloedige diere. Navorsing en ontdekkings sedert die 1970's dui egter daarop dat hulle aktiewe diere was met ’n verhoogde metabolisme wat verskeie aanpassings ondergaan het in sosiale interaksie.
Sedert die eerste dinosourusfossiel vroeg in die 19de eeu gevind is, is saamgestelde dinosourusskelette ’n gewilde trekpleister in museums dwarsoor die wêreld. Hulle het ’n plek in die wêreldkultuur en is gewild; as onderwerp in boeke en rolprente soos Jurassic Park of nuusitems wat dikwels in die media gedek word.
Etimologie
[wysig | wysig bron]Die takson Dinosauria is in 1842 amptelik so genoem deur die paleontoloog sir Richard Owen, wat dit gebruik het om te verwys na die "kenmerkende stam of suborde van souriese reptiele" wat in Engeland en dwarsoor die wêreld erken is.[4] Die term is afgelei van die Antieke Griekse woorde δεινός (deinos, "vreeslike of "verskriklik groot") en σαῦρος (sauros, "akkedis" of "reptiel").[4][5] Owen het dit bloot gebruik om te verwys na die grootte van die diere[6] en nie na hul voorkoms nie.
Definisie
[wysig | wysig bron]Volgens filogenetiese taksonomie word dinosourusse gewoonlik gedefinieer as ’n groep "bestaande uit Triceratops, Neornithes (moderne voëls), hul laaste gemeenskaplike voorsaat en alle afstammelinge".[7] Daar is ook al voorgestel dat Dinosauria gedefinieer word na aanleiding van die laaste gemeenskaplike voorsaat van Megalosaurus en Iguanodon, aangesien dit twee van die drie genera is wat Owen genoem het toe hy die Dinosauria erken het.[8] Albei definisies lei tot dieselfde groepe diere wat as dinosourusse beskou word.
- "Dinosauria = Ornithischia + Saurischia", en dit sluit in:
- Teropodes (meestal tweepotiges, karnivore en voëls)
- Ankilosouriërs (gepantserde, plantvretende vierpotiges)
- Stegosouriërs (plantvretende vierpotiges met plate)
- Seratopsiërs (plantvretende vierpotiges met horings en krae)
- Ornitopodes (twee- en vierpotige plantvreters, insluitende eendbekdinosourusse)
- Souropodomorfe (meestal groot, vierpotige plantvreters met lang nekke en sterte).[9]
Voëls word nou erken as die enigste oorlewende linie van teropodedinosourusse. In tradisionele taksonomie is voëls beskou as ’n aparte klas wat ontwikkel het uit dinosourusse, ’n uitgestorwe superorde. Die meeste moderne paleontoloë staan egter die idee voor dat vir ’n groep om natuurlik te wees, alle afstammelinge van lede van die groep ook in die groep ingesluit moet wees. Voëls word dus beskou as dinosourusse, en dinosourusse is dus nie uitgesterf nie. Voëls word geklassifiseer as behorende tot die subgroep Maniraptora, wat Coelurosauria is, wat Theropoda is, wat Saurischia is, wat dinosourusse is.[10]
Algemene beskrywing
[wysig | wysig bron]Dinosourusse kan in die algemeen beskryf word as argosourusse met ledemate wat reguit onder die lyf gehou word.[11] Baie prehistoriese diere word dikwels as dinosourusse beskou maar word nie wetenskaplik as dinosourusse geklassifiseer nie, soos ichtiosourusse, mosasourusse, plesiosourusse, pterosourusse en Dimetrodon; nie een van hulle het die reguit ledemate onder die lyf wat kenmerkend van ware dinosourusse is nie.[12]
Dinosourusse was die oorheersende landgewerweldes van die Mesosoïkum, veral die Jura- en Kryt-periode. Ander groepe diere het beperkte grootte en nisse gehad; soogdiere was byvoorbeeld selde groter as katte en was dikwels knaagdiergrootte-vreters van klein prooi.[13]
Dinosourusse was ’n groot groep uiteenlopende diere; volgens ’n 2006-studie is meer as 500 nie-vlieënde genera al tot dusver geïdentifiseer, en die totale getal genera in die fossielrekord word geraam op sowat 1 850, waarvan byna 75% nog ontdek moet word.[14] Volgens ’n vroeëre studie het daar sowat 3 400 dinosourusgenera bestaan, insluitende talle wat nie in die fossielrekord behoue sou gebly het nie.[15] Teen 17 September 2008 het altesaam 1 047 verskillende spesies dinosourusse al name gehad.[16] Van hulle was plantvreters, ander vleisvreters, insluitende saad-, vis-, insek- en allesvreters.
Terwyl die vroeë dinosourusse tweevoetig was (soos moderne voëls), was sommige prehistoriese spesies viervoetig en ander, soos Ammosaurus en Iguanodon, kon ewe maklik op twee of vier voete loop. Skedelaanpassings soos horings en plate was algemeen, terwyl sommige spesies pantsers gehad het. Hoewel hulle bekend is as groot diere, was die meeste dinosourusse in die Mesosoïkum so groot soos mense of kleiner. Dinosourusse was minstens teen die Vroeë Jura oor die wêreld versprei.[17] Moderne voëls bewoon feitlik alle habitats, van die land tot die see.
Geskiedenis van evolusie
[wysig | wysig bron]Oorsprong en vroeë evolusie
[wysig | wysig bron]
Dinosourusse het tydens die Middel- tot Laat Trias uit hul argosourus-voorsate ontwikkel, rofweg 20 miljoen jaar gelede nadat ’n geskatte 95% van alle lewe op aarde in die Perm-Trias-uitwissing uitgesterf het.[18][19] Radiometriese datering van die rotsformasie wat fossiele bevat van die vroeë dinosourus-genus Eoraptor van sowat 231,4 miljoen jaar gelede, wys dat hulle in dié tyd in die fossielrekord aanwesig was.[20] Paleontoloë dink Eoraptor verteenwoordig die gemeenskaplike voorsaat van alle dinosourusse;[21] as dit waar is, beteken dit die eerste dinosourusse was klein, tweevoetige roofdiere.[22] Die ontdekking van primitiewe, dinosourusagtige Ornithodira soos Marasuchus en Lagerpeton in Argentiniese Middel-Trias-strata ondersteun dié teorie; ontleding van fossiele dui daarop dat hierdie diere wel klein, tweevoetige roofdiere was. Dinosourusse kon al 243 miljoen jaar gelede hul verskyning gemaak het, soos gesien kan word uit oorblyfsels van die genus Nyasasaurus uit daardie periode, hoewel bekende fossiele te verbrokkel is om te bepaal of hulle dinosourusse was of net naby-verwante.[23]
Toe dinosourusse op die toneel verskyn, was die land bewoon deur verskeie soorte argosourusse en terapsides soos etosourusse, sinodonte, disinodonte en ornitosoegides. Die meeste van hierdie ander diere het uitgesterf in die Trias in een van twee uitwissingsvoorvalle. Die eerste was tussen die tydsnedes Karnium en Norium sowat 215 miljoen jaar gelede, toe disinodonte en ’n verskeidenheid argosouromorfe uitgesterf het. Dit is gevolg deur die Trias-Jura-uitwissing sowat 200 miljoen jaar gelede, wat die einde ingelui het vir die meeste van die ander groepe vroeë argosourusse soos die etosourusse, ornitosoegides en fitosourusse. Dié verliese het daartoe gelei dat die land oorheers is deur Krokodilagtiges, dinosourusse, soogdiere, pterosourusse en skilpaaie.[7] Die eerste paar linies van die vroeë dinosourusse het gediversifiseer in die Karnium- en die Norium-tydsnede van die Trias, moontlik deur die nisse te vul wat deur uitgestorwe spesies leeg gelaat is.[9]
Evolusie en paleobiogeografie
[wysig | wysig bron]Die evolusie van dinosourusse ná die Trias het gevolg op veranderings in plantegroei en die ligging van die kontinente. In die Laat Trias en Vroeë Jura was die landmassas verbind in een superkontinent, Pangea, en die wêreldwye dinosourusbevolkings het meestal bestaan uit selofisoïed-karnivore (vleisvreters) en vroeë souropodomorf-herbivore (plantvreters).[24] Naaksadiges (veral konifers), ’n moontlike voedselbron, het in die Laat Trias begin gedy. Vroeë souropodomorfe het nie gesofistikeerde meganismes gehad om kos in die mond te verwerk nie, en moes ander metodes verder af in die verteringskanaal gebruik om die kos op te breek.[25] Die algemene gelyksoortigheid van dinosourusse het tot in die Middel- en Laat Jura voortgeduur, met die meeste bevolkings wat bestaan het uit roofdiere soos seratosouriërs, spinosouroïede en karnosouriërs, en plantvreters soos stegosouriese ornitiskiums en groter souropodes. Ankilosouriërs en ornitopodes het ook al hoe algemener geraak, maar prosouropodes het uitgesterf. Konifers en pteridofiete was die mees algemene plante. Souropodes, soos die vroeëre prosouropodes, het ook nie kos in die mond verwerk nie, maar ornitiskiums het verskeie maniere ontwikkel om kos in die mond te hanteer, insluitende moontlike wang-agtige organe om die kos in die mond te hou, en kaakbewegings om dit fyn te maal.[25] Nog ’n noemenswaardige gebeurtenis in die Jura was die verskyning van ware voëls, wat afgestam het van selurosouriërs.[26]
Teen die Vroeë Kryt en die voortdurende verbrokkeling van Pangea, het dinosourusse op die onderskeie landmassas begin verskil. Vroeg in dié tydperk het ankilosouriërs, iguanodonte en brachiosourides deur Europa, Noord-Amerika en Noord-Afrika begin versprei. Hulle is later in Afrika aangevul of vervang deur groot spinosouride- en karcharodontosouride-teropodes, en rebbachisouride- en titanosouride-souropodes, wat ook in Suid-Amerika voorgekom het. In Asië het selurosouriërs soos dromeosourides, troödontides en oviraptorosouriërs algemene teropodes geword, en ankilosourides en vroeë seratopsiërs soos Psittacosaurus het belangrike plantvreters geword. Intussen het Australië die tuiste geword van primitiewe ankilosouriërs, hipsilofodonte en iguanodonte.[24] Dit lyk of die stegosouriërs in die een of ander stadium van die Vroeë Kryt en vroeë Laat Kryt uitgesterf het. ’n Groot verandering in die Vroeë Kryt, wat in die Laat Kryt in ’n groter mate voortgesit is, is die evolusie van blomplante. Terselfdertyd het verskeie groepe plantvreters meer gesofistikeerde maniere ontwikkel om kos in die mond met tande fyn te maal.
Daar was in die Laat Kryt drie groot bevolkings dinosourusse. Op die noordelike kontinente Noord-Amerika en Asië was die meeste teropodes tirannosourides en verskeie soorte kleiner teropodes, terwyl die plantvreters veral bestaan het uit hadrosourides, seratopsiërs, ankilosourides en pachisefalosouriërs. Op die suidelike kontinente, wat die nou verbrokkelende Gondwana uitgemaak het, was abelisourides die algemene teropodes, en titanosouriër-souropodes die algemene plantvreters. In Europa was dromeosourides, rabdodontide-iguanodonte, nodosouride-ankilosouriërs en titanosouriër-souropodes volop.[24] Blomplante het toegeneem[25] en die eerste grassoorte het aan die einde van die Kryt verskyn.[27] Hadrosourides, wat kos gemaal het, en seratopsiërs, wat kos stukkend geskeur het, het wydverspreid in Noord-Amerika en Asië geword. Teropodes het ook versprei as plant- en allesvreters, en terisinosouriërs en ornitomimosouriërs het algemeen geraak.[25]
In die Kryt-Paleogeen-uitwissing van sowat 66 miljoen jaar gelede, aan die einde van die Kryt-periode, het alle dinosourusgroepe uitgesterf, behalwe Neornithes-voëls. Sommige ander diapsidegroepe soos Krokodilagtiges, sebesosoegiërs, skilpaaie, akkedisse, slange, sfenodontes en choristoderas het ook oorleef.[28]
Die oorlewende linies Neornithes-voëls, insluitende die voorsate van moderne loopvoëls (eende en hoenders) en watervoëls, het aan die begin van die Paleogeen-periode vinning gediversifiseer en nisse gevul wat leeg gelaat is deur dinosourusgroepe. Soogdiere het egter ook vinnig gediversifiseer en het die nisse op land uiteindelik oorheers.[29]
Klassifikasie
[wysig | wysig bron]- Hoofartikel: Klassifikasie van dinosourusse
Dinosourusse is argosourusse, nes moderne krokodilagtiges. Binne die argosourusgroep word dinosourusse veral deur hul loop onderskei. Dinosourusse se bene strek van die lyf af ondertoe, terwyl die bene van akkedisse en krokodille na die kant toe uitstrek.[30]
Dinosourusse as ’n klade word verdeel in twee hoofgroepe: souriskiums (Saurischia) en ornitiskiums (Ornithischia). Souriskiums sluit in dié taksa wat ’n meer onlangse gemeenskaplike voorsaat met voëls deel as met ornitiskiums, terwyl ornitiskiums dié taksa insluit wat ’n meer onlangse gemeenskaplike voorsaat met Triceratops deel as met souriskiums. Anatomies kan die twee groepe veral uitmekaar geken word deur die struktuur van hul pelvis. Vroeë souriskiums ("akkedisheupe", van die Griekse σαῦρος sauros, "akkedis", en ἰσχίον ischion, heupgewrig) het die heupstruktuur van hul voorsate behou. Ornitiskiums ("voëlheupe", van die Griekse ορνιθειος ornitheos, "voël", en ισχιον ischion, "heupgewrig") het ’n aangepaste pelvisstruktuur.
-
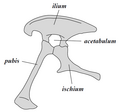 Saurischia-pelvisstruktuur (linkerkant).
Saurischia-pelvisstruktuur (linkerkant). -
 Tirannosourus-pelvis (wys die Saurischia-struktuur – linkerkant).
Tirannosourus-pelvis (wys die Saurischia-struktuur – linkerkant). -
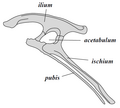 Ornithischia-pelvisstruktuur (linkerkant).
Ornithischia-pelvisstruktuur (linkerkant). -
 Edmontosourus-pelvis (wys die Ornithischia-struktuur – linkerkant)
Edmontosourus-pelvis (wys die Ornithischia-struktuur – linkerkant)




Taksonomie
[wysig | wysig bron]


Die volgende lys is ’n vereenvoudigde klassifikasie van groepe gebaseer op hul evolusionêre verwantskappe. Dit is georganiseer volgens ’n lys Mesosoïese dinosourusspesies deur Holtz (2008).[31]
Die dolk (†) toon groepe aan met geen oorlewende lede nie.
- Dinosauria
- Saurischia ("met akkedisheupe"; sluit in Theropoda en Sauropodomorpha)
- Theropoda (alle tweevoetiges; die meeste was karnivore)
- †Herrerasauria (vroeë tweevoetige karnivore)
- †Coelophysoidae (klein, vroeë teropodes; sluit in Coelophysis en naby-verwante)
- †Dilophosauridae (vroeë vleisetende teropodes met kuiwe)
- †Ceratosauria (met horings)
- Tetanurae ("met reguit sterte"; sluit in die meeste teropodes)
- †Megalosauroidea (vroeë groep groot karnivore)
- †Carnosauria (Allosaurus en naby-verwante soos Carcharodontosaurus)
- Coelurosauria (geveerde teropodes)
- †Compsognathidae (algemene vroeë Coelurosauria met klein voorste ledemate)
- †Tyrannosauridae (Tyrannosaurus en naby-verwante met klein voorste ledemate)
- †Ornithomimosauria ("soos volstruise"; meestal sonder tande; karnivore tot moontlik herbivore)
- †Alvarezsauroidea (klein insektivore met klein voorste ledemate en een groot klou)
- Maniraptora (met lang, skraal arms en vingers)
- †Therizinosauria (tweevoetige herbivore met groot kloue en klein koppe)
- †Oviraptorosauria (meestal sonder tande)
- †Archaeopterygidae (klein teropodes met vlerke, of primitiewe voëls)
- †Deinonychosauria (klein tot mediumgrootte en voëlagtig met ’n kenmerkende toonklou)
- Avialae (moderne voëls en hul naby-verwante)
- †Scansoriopterygidae (klein, primitiewe voëls met lang derde vingers)
- †Omnivoropterygidae (groot, vroeë voëls met kort sterte)
- †Confuciusornithidae (klein, tandlose voëls)
- †Enantiornithes (primitiewe, vlieënde voëls wat in bome gehou het)
- Euornithes (gevorderde vlieënde voëls)
- †Yanornithiformes (Sjinese voëls uit die Kryt met tande)
- †Hesperornithes (duikende watervoëls)
- Aves (moderne voëls met snawels en hul uitgestorwe verwante)

- †Sauropodomorpha (herbivore met klein koppe en lang nekke en sterte)
- †Guaibasauridae (klein, primitiewe Sauropodomorpha-omnivore)
- †Plateosauridae (primitiewe, tweevoetige Prosauropoda)
- †Riojasauridae (klein, primitiewe Sauropodomorpha)
- †Massospondylidae (klein, primitiewe Sauropodomorpha)
- †Sauropoda (groot en swaar viervoetiges)
- †Vulcanodontidae (primitiewe Sauropoda met pilaaragtige ledemate)
- †Eusauropoda ("ware Sauropoda")
- †Cetiosauridae ("walvis-reptiele")
- †Turiasauria (Europese groep Jura- en Kryt-Sauropoda)
- †Neosauropoda ("nuwe Sauropoda")
- †Diplodocoidea (lang skedels en sterte; tande gewoonlik smal)
- †Macronaria (boksagtige skedels; lepel- of potloodvormige tande)
- †Brachiosauridae (met lang nekke en lang arms)
- †Titanosauria (uiteenlopend; stewig met breë heupe)

- †Ornithischia ("met voëlheupe"; uiteenlopende twee- en viervoetige herbivore)
- †Heterodontosauridae (klein herbi-/omnivoor-ornitopodes met prominente snytande)
- †Thyreophora (gepantserde dinosourusse; meestal viervoetig)
- †Ankylosauria (met skildplate as primêre pantsers; sommige met knuppelagtige sterte)
- †Stegosauria (skerp punte en plate as primêre pantsers)
- †Neornithischia ("nuwe Ornithischia")
- †Ornithopoda (verskeie groottes; twee- en viervoetiges; talle met tande en ’n kenmerkende manier van kou)
- †Marginocephalia (gekenmerk deur kopbeengroei)
- †Pachycephalosauria (tweevoetiges met knobbelrige groeisels aan skedels)
- †Ceratopsia (viervoetiges met krae; sommige met horings)
Biologie
[wysig | wysig bron]
Kennis oor dinosourusse kom van ’n verskeidenheid fossiele en niefossiele rekords, insluitende gefossileerde bene, uitwerpsels, vere, velafdrukke, interne organe, gastroliete, voetspore en sagte weefsel.[32][33] Baie studieterreine dra by tot die mens se begrip van dinosourusse, insluitende fisika, chemie, biologie en paleontologie. Twee onderwerpe wat veral deeglik bestudeer word, is grootte en gedrag.[34]
Grootte
[wysig | wysig bron]Bewyse dui daarop dat die grootte van dinosourusse gewissel het tydens die geologiese periodes Trias, Vroeë Jura, Laat Jura en Kryt.[21] Teropode-roofdiere in die Mesosoïkum-era het gewoonlik tussen 100 en 1 000 kg geweeg, terwyl huidige soogroofdiere hoogstens 10 tot 100 kg weeg.[35]
Die souropodes was die grootste en swaarste dinosourusse. Vir ’n groot deel van die dinosourus-tydperk was die kleinste souropodes groter as enigiets anders in hul habitat, en die grootste een was ’n grootteorde groter as enigiets anders wat sedertdien die aarde bewandel het. Reusagtige prehistoriese soogdiere soos die Paraceratherium (die grootste landsoogdier nog) is verdwerg deur die groot souropodes, en net moderne walvisse is byna so groot of groter.[36]
Grootste en kleinste
[wysig | wysig bron]

Wetenskaplikes sal waarskynlik nooit seker wees watter dinosourusse die grootste of die kleinste was wat ooit bestaan het nie. Dit is omdat net ’n baie klein persentasie diere gefossileer word, en die meeste daarvan is onder die grond begrawe. Min van die fossiele wat ontdek word, is volledige skelette, en afdrukke van velle en ander sagte weefsel is skaars. Die rekonstruksie van volledige skelette deur ’n vergelyking met die grootte en morfologie van soortgelyke, beter bekende spesies is ’n onnoukeurige kuns, en die rekonstruksie van spiere en ander organe is hoogstens ingeligte raaiskote.[37]
Die langste en swaarste dinosourus volgens goeie fossiele is Giraffatitan brancai (voorheen geklassifiseer as ’n spesie van Brachiosaurus). Die oorblyfsels is tussen 1907 en 1912 in Tanzanië ontdek. Die bene van verskeie individue van dieselfde grootte is in die skelet geïnkorporeer;[38] dit is 12 m hoog en 22,5 m lank, en sou behoort het aan ’n dier wat tussen 30 000 en 60 000 kg geweeg het. Die langste volledige dinosourus is die 27 m lange Diplodocus, wat in 1907 in Wyoming, VSA, ontdek is.[39]
Daar was groter dinosourusse, maar kennis oor hulle is gebaseer op ’n paar stukke fossiele. Die meeste van die grootste plantvretende voorbeelde is almal in die 1970's of later ontdek, en sluit in die massiewe Argentinosaurus, wat tussen 80 000 en 100 000 kg kon geweeg het; van die langstes was die 33,5 m lange Diplodocus hallorum[40] en die 33 m lange Supersaurus.[41] Die hoogste een was die 18 m hoë Sauroposeidon, so hoog soos ’n gebou van ses verdiepings. Die swaarste en langste van hulle almal was dalk Amphicoelias fragillimus, wat net bekend is van een, nou verlore, gedeeltelike werwelboog wat in 1878 beskryf is. Die dier kon 58 m lank gewees en meer as 120 000 kg geweeg het.[40] Die grootste bekende karnivoor was Spinosaurus, wat 16 tot 18 m lank was en 8 150 kg kon geweeg het.[42]
As ’n mens nie voëls insluit nie, was die kleinste bekende dinosourus omtrent so groot soos ’n duif.[43] Die kleinste nie-vlieënde dinosourusse was die naaste aan voëls verwant. Die algehele lengte van Anchiornis huxleyi se skelet was byvoorbeeld minder as 35 cm.[43][44] A. huxleyi is tans die kleinste nie-vlieënde dinosourus wat van ’n volwasse fossiel beskryf is, met ’n geskatte gewig van 110 g.[44] Die kleinste plantvretende, nie-vlieënde dinosourus sluit in Microceratus en Wannanosaurus (elk sowat 60 cm lank).[31][45]
Gedrag
[wysig | wysig bron]
Baie moderne voëls is baie sosiaal en kom dikwels in swerms voor. Daar is algemene konsensus dat ’n deel van die gedrag van voëls, sowel as krokodille (voëls se naaste lewende verwante), ook algemeen in uitgestorwe dinosourusse was. Vertolkings van die gedrag van fossiele spesies word gewoonlik gebaseer op die posisie van skelette en hul habitat, rekenaarsimulasies van hul biomeganika en vergelykings met moderne diere in soortgelyke ekologiese nisse.
Die eerste moontlike bewyse van groepvorming as ’n algemene gedrag onder dinosourusse buiten voëls was die ontdekking in 1878 van 31 Iguanodon bernissartensis in België. Daar is destyds geglo die ornitiskiums het omgekom nadat hulle saam in ’n diep sinkgat geval en verdrink het.[46] Ander voorbeelde van groepe diere wat saam gesterf het, is sedertdien ontdek. Dit, tesame met die spore van groepe diere, dui daarop dat groepvorming algemeen was onder baie vroeë dinosourusspesies. Voetspore van honderde of selfs duisende herbivore dui daarop dat hulle in groot troppe beweeg het, soos bisons of springbokke. Souropode-spore bewys dié diere het beweeg in groepe wat uit meer as een spesie bestaan het, of minstens in Oxfordshire, Engeland,[47] hoewel daar geen ander bewyse is van troppe nie.[48]
Om saam in groepe te beweeg, was dalk ’n metode van verdediging, vir migrasiedoeleindes of om beskerming aan die jong diere te bied. Daar is bewyse dat baie soorte stadig groeiende dinosourusse, insluitende verskeie teropodes, souropodes, ankilosouriërs, ornitopodes en seratopsiërs, groepe van onvolwasse individue gevorm het. Daar word ook geglo dinosourusse het troppe gevorm om saam groot prooi te jag.[49] Dié gedrag is egter ongewoon onder voëls, krokodille en ander reptiele.

Uit ’n gedragsoogpunt is een van die waardevolste fossiele in 1971 in die Gobiwoestyn ontdek. Dit het ’n velosiraptor ingesluit wat ’n protoseratops aanval;[50] dit het ’n bewys verskaf dat dinosourusse mekaar wel aangeval het.[51] Nog ’n bewys is die fossiel van ’n gedeeltelik herstelde stert van ’n edmontosourus; die stert is op so ’n manier beskadig dat dit wys die dier is gebyt deur ’n tirannosourus, maar dat dit die aanval oorleef het.[51]
Vergelykings tussen die buiteoogvliesringe van dinosourusse en moderne voëls is al gebruik om die aktiwiteitspatrone van dinosourusse vas te stel. Hoewel sommige wetenskaplikes meen dinosourusse was bedags aktief, wys hierdie vergelykings klein roofdiere soos die dromeosourides, juravenators en megapnosourusse was waarskynlik snags aktief. Groot en middelslag-dinosourusse kon bedags en snags ewe aktief gewees het, hoewel die klein ornitiskium agilisourus bedags aktief was.[52]
Kommunikasie
[wysig | wysig bron]Dit is bekend dat moderne voëls kommunikeer deur middel van visuele en klankseine, en die verskeidenheid van visuele vertoonstrukture onder fossiele dinosourusgroepe toon aan visuele kommunikasie was belangrik onder hulle. Oor die gebruik van klank is egter minder bekend. In 2008 het die paleontoloog Phil Senter bevind die meeste Mesosoïese dinosourusse het nie geluide gemaak nie, ondanks die uitbeelding van brullende dinosourusse in rolprente.[53][54] Hy het bevind die stembande in die larinks het waarskynlik verskeie kere ontwikkel in reptiele, insluitende krokodille, wat gromgeluide in die keel kan maak. Voëls het egter nie ’n larinks nie en gebruik ’n sirinks, ’n stemorgaan wat uniek is aan voëls.[55]
Uitwissing van groot groepe
[wysig | wysig bron]- Hoofartikel: Kryt-Paleogeen-uitwissing

Die ontdekking dat voëls ’n soort dinosourus is, wys die dinosourusbevolking as ’n geheel het nie uitgesterf soos meestal gemeen word nie.[56] Alle nie-vlieënde dinosourusse en baie groepe voëls het egter wel sowat 66 miljoen jaar gelede uitgesterf tydens die Kryt-Paleogeen-uitwissing. Baie ander groepe diere het ook in dié tyd uitgesterf, onder meer ammoniete, plesiosourusse, pterosourusse en baie groepe soogdiere.[17] Daar was nie groot verliese onder insekte nie en hulle was dus waarskynlik kos vir van die oorlewende dierspesies.
Die aard van die massauitwissing word sedert die 1970's baie goed nagevors. Daar is tans verskeie teorieë daaroor en die een wat deur die meeste wetenskaplikes ondersteun word, is dat dit veroorsaak is deur ’n groot komeet of asteroïde wat die aarde getref het en ’n katastrofale uitwerking op die omgewing gehad het.
Chicxulub-krater
[wysig | wysig bron]Die impakteorie is ondersteun deur die ontdekking van die 18 km wye Chicxulub-krater in die laat 1970's in die Golf van Meksiko,[57] wat beskou word as bewys van ’n groot, katastrofale impak.[58] As en stof het daarna onder meer die son se strale afgekeer en dus fotosintese onmoontlik gemaak.[59] Brande het ook waarskynlik wêreldwyd voorgekom weens die hitte en brandende brokstukke wat deur die impak veroorsaak is. Onlangse navorsing het getoon die wêreldwye puinlaag wat deur die impak gevorm is, bevat genoeg roet om te kan aflei die hele biosfeer het gebrand.[60]
Die asteroïde (of komeet) het in die see geland en sou enorme tsoenami's veroorsaak het, waarvoor bewyse gevind is op verskeie plekke in die Oos-VSA en om die Karibiese See – seesand op plekke wat toe in die binneland geleë was, en plantoorblyfsels en aardrotse in see-sedimente wat dateer uit die tyd van die impak.
In Maart 2010 het ’n internasionale paneel wetenskaplikes die asteroïde-teorie, spesifiek die Chicxulub-impak, onderskryf as die oorsaak van die uitwissing. ’n Span van 41 wetenskaplikes het 20 jaar se wetenskaplike geskrifte nagevors en enige teorieë oor enorme vulkaniese uitbarstings verwerp. Hulle het vasgestel die uitwissing is veroorsaak deur die Chicxulub-impak, wat soveel energie opgewek het as 100 teraton TNT – meer as ’n miljard keer die energie van die bomme op Nagasaki en Hirosjima.[58]
Daar is egter wetenskaplikes wat glo die uitwissing is veroorsaak of vererger deur ander faktore soos vulkaniese uitbarstings,[61] klimaatsveranderings en/of ’n verandering in seevlakke.
Dekan-trappe
[wysig | wysig bron]
Voor 2000 is argumente dat basaltlawa van die Dekan-trappe in Indië die uitwissing veroorsaak het, gewoonlik verbind met die opinie dat die uitwissing geleidelik plaasgevind het. Basaltlawa-voorvalle het omstreeks 68 miljoen jaar gelede begin en langer as 2 miljoen jaar geduur. Die mees onlangse getuienis dui daarop dat die trappe oor ’n tydperk van 800 000 jaar plaasgevind het, oor die tyd van die Kryt-Paleogeen-grens (K-Pg-grens), en dit kon verantwoordelik gewees het vir die uitwissing en die stadige herstel van die biotika daarna.[62]
Regressie
[wysig | wysig bron]Daar is duidelike bewyse dat seevlakke in die Maastricht, die laaste etage van die Kryt-periode, meer gedaal het as op enige ander tyd in die Mesosoïese Era. Dit is moontlik veroorsaak deurdat oseaanriwwe minder aktief geraak en toe onder hul eie gewig gesink het.[17][63]
’n Groot regressie sou die gebied van die kontinentale plat aansienlik verklein het – en dit is waar die meeste seespesies aangetref is. Die regressie sou ook klimaatsveranderings teweeggebring het, deels deur winde en seestrome te ontwrig en deels deur die Aarde se albedo te verminder en temperature wêreldwyd te laat toeneem.[64]
Regressie het ook die verlies aan binneseë tot gevolg gehad. Dit het habitats in ’n groot mate verander en kusvlaktes vernietig.
Moontlike oorlewende dinosourusse
[wysig | wysig bron]Die fossiele van nie-vlieënde dinosourusse word soms bo die K-Pg-grens ontdek. In 2001 het die paleontoloë Zielinski en Budahn na bewering ’n enkele Hadrosauridae-beenfossiel in die San Juan-vallei in Nieu-Mexiko ontdek en dit beskryf as ’n bewys dat dinosourusse in die Paleoseen-epog voorgekom het. Die formasie waarin dit ontdek is, is gedateer as komende uit die Paleoseen, sowat 64,5 miljoen jaar gelede. As die been nie vanweë weerstoestande na ’n jonger stratum verplaas is nie, sou dit wel ’n bewys wees dat sommige dinosourus-bevolkings vir minstens ’n halfmiljoen jaar in die Kainosoïese Era oorleef het.[65]
Nog bewyse sluit in dinosourus-oorblyfsels in die Hell Creek-formasie in Amerika wat 40 000 jaar jonger as die K-Pg-grens is. Ook in Sjina en ander dele van die wêreld was daar berigte oor sulke vondste.[66] Baie wetenskaplikes glo die bene het uitgespoel en is toe in veel jonger afsettings herbegrawe.[67][68] Direkte datering van die bene self steun egter die jonger datum, en ander daterings dui op ’n ouderdom van 64,8 miljoen jaar.[69]
Al is dit korrek, weerspreek die vonds van ’n paar dinosourusse wat tot in die vroeë Paleoseen oorleef het, nie die feite van die massauitwissing nie.[67]
Verwysings
[wysig | wysig bron]- ↑ Gauthier, Jacques; de Querioz, Kevin. "Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name 'Aves'.". New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom. Peabody Museum of Natural History, Yale University. ISBN 0-912532-57-2.
{{cite book}}:|access-date=requires|url=(hulp); Onbekende parameter|chapterurl=geïgnoreer (hulp) - ↑ Wang, S.C., and Dodson, P. (2006). "Estimating the Diversity of Dinosaurs". Proceedings of the National Academy of Sciences USA. 103 (37): 13601–13605. doi:10.1073/pnas.0606028103. PMID 16954187.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Will the real dinosaurs stand up?, BBC, September 17, 2008
- ↑ 4,0 4,1 Owen, R (1842). Report on British Fossil Reptiles." Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841. London: John Murray. pp. 60–204.
- ↑ "Liddell–Scott–Jones Lexicon of Classical Greek". Besoek op 5 Augustus 2008.
- ↑ Farlow, J.O., and Brett-Surman, M.K. (1997). "Preface". In Farlow, J.O., and Brett-Surman, M.K. (reds.) (red.). The Complete Dinosaur. Indiana University Press. pp. ix–xi. ISBN 0-253-33349-0.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ 7,0 7,1 Benton, Michael J. (2004). "Origin and relationships of Dinosauria". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (reds.) (red.). The Dinosauria (2nd uitg.). Berkeley: University of California Press. pp. 7–19. ISBN 0-520-24209-2.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ Olshevsky, G. (2000). "An annotated checklist of dinosaur species by continent". Mesozoic Meanderings. 3: 1–157.
- ↑ 9,0 9,1 Langer, Max C.; Martin D. Ezcurra; Jonathas S. Bittencourt; Fernando E. Novas (2010). "The origin and early evolution of dinosaurs". Biological Reviews. 85 (1): 65–66, 82. doi:10.1111/j.1469-185x.2009.00094.x. PMID 19895605.
- ↑ Padian, K. (2004). "Basal Avialae". In Weishampel, D.B.; Dodson, P.; Osmolska, H. (reds.). The Dinosauria (2de uitg.). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2
- ↑ Glut, Donald F. (1997). Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. p. 40. ISBN 0-89950-917-7.
- ↑ Lambert, David; the Diagram Group (1990). The Dinosaur Data Book. New York: Avon Books. p. 288. ISBN 978-0-380-75896-8.
- ↑ Morales, Michael (1997). "Nondinosaurian vertebrates of the Mesozoic". In Farlow JO, Brett-Surman MK (red.). The Complete Dinosaur. Bloomington: Indiana University Press. pp. 607–624. ISBN 0-253-33349-0.
- ↑ Wang, S.C., and Dodson, P. (2006). "Estimating the Diversity of Dinosaurs". Proceedings of the National Academy of Sciences USA. 103 (37): 13601–13605. Bibcode:2006PNAS..10313601W. doi:10.1073/pnas.0606028103. PMC 1564218. PMID 16954187.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Russell, Dale A. (1995). "China and the lost worlds of the dinosaurian era". Historical Biology. 10: 3–12. doi:10.1080/10292389509380510.
- ↑ Amos J (17 September 2008). "Will the real dinosaurs stand up?". BBC News (in Engels). Geargiveer vanaf die oorspronklike op 4 Junie 2019. Besoek op 23 Maart 2011.
- ↑ 17,0 17,1 17,2 MacLeod, N, Rawson, PF, Forey, PL, Banner, FT, Boudagher-Fadel, MK, Bown, PR, Burnett, JA, Chambers, P, Culver, S, Evans, SE, Jeffery, C, Kaminski, MA, Lord, AR, Milner, AC, Milner, AR, Morris, N, Owen, E, Rosen, BR, Smith, AB, Taylor, PD, Urquhart, E & Young, JR (1997). "The Cretaceous–Tertiary biotic transition". Journal of the Geological Society. 154 (2): 265–292. doi:10.1144/gsjgs.154.2.0265.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Kump LR, Pavlov A & Arthur MA (2005). "Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia". Geology. 33 (5): 397–400. Bibcode:2005Geo....33..397K. doi:10.1130/G21295.1.
- ↑ Tanner LH, Lucas SG & Chapman MG (2004). "Assessing the record and causes of Late Triassic extinctions" (PDF). Earth-Science Reviews. 65 (1–2): 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Geargiveer vanaf die oorspronklike (PDF) op 25 Oktober 2007. Besoek op 22 Oktober 2007.
- ↑ Alcober, Oscar A.; Martinez, Ricardo N. (2010). "A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina". ZooKeys. 63 (63): 55–81. doi:10.3897/zookeys.63.550. PMC 3088398. PMID 21594020. [1]
- ↑ 21,0 21,1 Sereno PC (1999). "The evolution of dinosaurs". Science. 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. PMID 10381873.
- ↑ Sereno, P.C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria". Nature. 361 (6407): 64–66. Bibcode:1993Natur.361...64S. doi:10.1038/361064a0.
- ↑ Nesbitt, S. J., Barrett, P. M., Werning, S., Sidor, C. A. en A. J. Charig. (2012). "The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania." Biology Letters.
- ↑ 24,0 24,1 24,2 Holtz, Thomas R. Jr.; Chapman, Ralph E.; Lamanna, Matthew C. (2004). "Mesozoic biogeography of Dinosauria". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (reds.). The Dinosauria (2nd uitg.). Berkeley: University of California Press. pp. 627–642. ISBN 978-0-520-24209-8.
- ↑ 25,0 25,1 25,2 25,3 Fastovsky, David E.; Smith, Joshua B. (2004). "Dinosaur paleoecology". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (reds.). The Dinosauria (2nd uitg.). Berkeley: University of California Press. pp. 614–626. ISBN 978-0-520-24209-8.
- ↑ Padian K (2004). "Basal avialae". In Weishampel DB, Dodson P, Osmólska H (red.). The Dinosauria (2d edition). University of California Press. pp. 210–231. ISBN 0-520-24209-2.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ Prasad, V.; Strömberg, CA; Alimohammadian, H; Sahni, A (2005). "Dinosaur coprolites and the early evolution of grasses and grazers". Science. 310 (5751): 1170–1180. Bibcode:2005Sci...310.1177P. doi:10.1126/science.1118806. PMID 16293759.
- ↑ Archibald, J. David; Fastovsky, David E. (2004). "Dinosaur Extinction". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (reds.). The Dinosauria (2nd uitg.). Berkeley: University of California Press. pp. 672–684. ISBN 978-0-520-24209-8.
- ↑ Lindow, B.E.K. (2011). "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification." In Dyke, G. en Kaiser, G. (reds.)Living Dinosaurs: The Evolutionary History of Modern Birds, John Wiley & Sons, Ltd, Chichester, UK. doi:10.1002/9781119990475.ch14
- ↑ Brusatte, Stephen L. (2012). Dinosaur Paleobiology (1. uitg.). New York: Wiley, J. pp. 9–20, 21. ISBN 978-0-470-65658-7.
- ↑ 31,0 31,1 Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- ↑ Dal Sasso, C. and Signore, M. (1998). "Exceptional soft-tissue preservation in a theropod dinosaur from Italy". Nature. 392 (6674): 383–387. Bibcode:1998Natur.392..383D. doi:10.1038/32884.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. (2005). "Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex". Science. 307 (5717): 1952–1955. Bibcode:2005Sci...307.1952S. doi:10.1126/science.1108397. PMID 15790853.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Weishampel, D.B., Dodson, P., Oslmolska, H. (1990). "The Dinosauria". University of California Press. pp. 733. ISBN 0-520-06727-4
- ↑ Farlow JA (1993). "On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs". Functional Morphology and Evolution. American Journal of Science, Special Volume 293-A. pp. 167–199.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ Sander, P. Martin; Christian, Andreas; Clauss, Marcus; Fechner, Regina; Gee, Carole T.; Griebeler, Eva-Maria; Gunga, Hanns-Christian; Hummel, Jürgen; Mallison, Heinrich (2011). "Biology of the sauropod dinosaurs: the evolution of gigantism". Biological Reviews. 86 (1): 117–155. doi:10.1111/j.1469-185X.2010.00137.x. PMC 3045712. PMID 21251189.
{{cite journal}}: Onbekende parameter|displayauthors=geïgnoreer (hulp) - ↑ Paul, Gregory S. (2010). Princeton Field Guide to Dinosaurs. Princeton University Press. ISBN 978-0-691-13720-9.
- ↑ Colbert, Edwin Harris (1971). Men and dinosaurs: the search in field and laboratory. Harmondsworth [Eng.]: Penguin. ISBN 0-14-021288-4.
- ↑ Lucas, H., Hecket, H. & S. (2004). "Reappraisal of Seismosaurus, a Late Jurassic Sauropod". Proceeding, Annual Meeting of the Society of Paleontology 36 (5): 422.
- ↑ 40,0 40,1 Carpenter, Kenneth (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus". In Foster, John R.; and Lucas, Spencer G. (reds.) (red.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque: New Mexico Museum of Natural History and Science. pp. 131–138.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ Lovelace, David M. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional. 65 (4): 527–544.
- ↑ dal Sasso C, Maganuco S, Buffetaut E, Mendez MA (2006). "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities" (PDF). Journal of Vertebrate Paleontology. 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. Geargiveer vanaf die oorspronklike (PDF) op 29 April 2011. Besoek op 5 Mei 2011.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ 43,0 43,1 Zhang F, Zhou Z, Xu X, Wang X, Sullivan C (2008). "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers". Nature. 455 (7216): 1105–1108. Bibcode:2008Natur.455.1105Z. doi:10.1038/nature07447. PMID 18948955.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ 44,0 44,1 Xu X, Zhao Q, Norell M, Sullivan C, Hone D, Erickson G, Wang XL, Han FL, Guo Y (2008). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin". Chinese Science Bulletin. 54 (3): 430–435. doi:10.1007/s11434-009-0009-6.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Butler, R.J.; Zhao, Q. (2009). "The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China". Cretaceous Research. 30 (1): 63–77. doi:10.1016/j.cretres.2008.03.002.
- ↑ Yans J, Dejax J, Pons D, Dupuis C & Taquet P (2005). "Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)". Comptes Rendus Palevol (in French). 4 (1–2): 135–150. doi:10.1016/j.crpv.2004.12.003.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) AS1-onderhoud: onerkende taal (link) - ↑ Day, J.J.; Upchurch, P; Norman, DB; Gale, AS; Powell, HP (2002). "Sauropod trackways, evolution, and behavior". Science. 296 (5573): 1659. doi:10.1126/science.1070167. PMID 12040187.
- ↑ Wright, Joanna L. (2005). "Steps in understanding sauropod biology". In Curry Rogers, Kristina A.; and Wilson, Jeffrey A. (red.). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 252–284. ISBN 0-520-24623-3.
{{cite book}}: AS1-onderhoud: meer as een naam (link) - ↑ Lessem, Don; Glut, Donald F. (1993). "Allosaurus". The Dinosaur Society's Dinosaur Encyclopedia. Random House. pp. 19–20. ISBN 978-0-679-41770-5.
- ↑ "The Fighting Dinosaurs" (in Engels). American Museum of Natural History. Geargiveer vanaf die oorspronklike op 9 Maart 2012. Besoek op 5 Desember 2007.
- ↑ 51,0 51,1 Carpenter, K. (1998). "Evidence of predatory behavior by theropod dinosaurs". Gaia. 15: 135–144.[dooie skakel]
- ↑ Schmitz, L.; Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030): 705–708. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Hopson, James A. (1975). "The evolution of cranial display structures in hadrosaurian dinosaurs". Paleobiology. 1 (1): 21–43.
- ↑ Diegert, Carl F. (1998). "A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen". Journal of Vertebrate Paleontology. 18 (3, Suppl.): 38A.
- ↑ Senter, P. (2008). "Voices of the past: a review of Paleozoic and Mesozoic animal sounds". Historical Biology. 20 (4): 255–287. doi:10.1080/08912960903033327.
- ↑ Dingus, L. and Rowe, T. (1998). The Mistaken Extinction – Dinosaur Evolution and the Origin of Birds. New York: W. H. Freeman.
- ↑ Hildebrand, A. R., G. T. Penfield, et al. (1991). "Chicxulub crater: a possible Cretaceous/Tertiary boundary impact crater on the Yucatan peninsula, Mexico." Geology 19: 867-871.
- ↑ 58,0 58,1 Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J. A.; Barton, P. J.; Bown, P. R.; Bralower, T. J.; Christeson, G. L.; Claeys, P.; Cockell, C. S.; Collins, G. S.; Deutsch, A.; Goldin, T. J.; Goto, K.; Grajales-Nishimura, J. M.; Grieve, R. A. F.; Gulick, S. P. S.; Johnson, K. R.; Kiessling, W.; Koeberl, C.; Kring, D. A.; MacLeod, K. G.; Matsui, T.; Melosh, J.; Montanari, A.; Morgan, J. V.; Neal, C. R.; Nichols, D. J.; Norris, R. D.; Pierazzo, E. (5 Maart 2010). "The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous- Paleogene Boundary |". Science 327 (5970): 1214–1218. Bibcode:2010Sci...327.1214S.doi:10.1126/science.1177265
- ↑ Alvarez LW, Alvarez W, Asaro F, Michel HV (1980). "Extraterrestrial cause for the Cretaceous–Tertiary extinction". Science. 208 (4448): 1095–1108. Bibcode:1980Sci...208.1095A. doi:10.1126/science.208.4448.1095.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Robertson, D.S., Lewis, W.M., Sheehan, P.M. & Toon, O.B. (2013) K/Pg Extinction: Re-evaluation of the Heat/Fire Hypothesis.Journal of Geophysical Research: Biogeosciences
- ↑ Keller, G. (2012). The Cretaceous--Tertiary Mass Extinction, Chicxulub Impact, and Deccan Volcanism. Earth and Life, Springer: 759--793.
- ↑ Keller G, Adatte T, Gardin S, Bartolini A, Bajpai S (2008). "Main Deccan volcanism phase ends near the K–T boundary: Evidence from the Krishna-Godavari Basin, SE India". Earth and Planetary Science Letters. 268 (3–4): 293–311. Bibcode:2008E&PSL.268..293K. doi:10.1016/j.epsl.2008.01.015.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Li, Liangquan; Keller, Gerta (1998). "Abrupt deep-sea warming at the end of the Cretaceous". Geology. 26 (11): 995–998. Bibcode:1998Geo....26..995L. doi:10.1130/0091-7613(1998)026<0995:ADSWAT>2.3.CO;2.
- ↑ Marshall CR, Ward PD (1996). "Sudden and Gradual Molluscan Extinctions in the Latest Cretaceous of Western European Tethys". Science. 274 (5291): 1360–1363. Bibcode:1996Sci...274.1360M. doi:10.1126/science.274.5291.1360. PMID 8910273.
- ↑ Fassett, JE, Lucas, SG, Zielinski, RA, and Budahn, JR; Lucas; Zielinski; Budahn (2001). "Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA" (PDF). Catastrophic events and mass extinctions, Lunar and Planetary Contribution. 1053: 45–46. Bibcode:2001caev.conf.3139F. Besoek op 18 Mei 2007.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Sloan, R. E., Rigby, K,. Van Valen, L. M., Gabriel, Diane (1986). "Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek Formation". Science. 232 (4750): 629–633. Bibcode:1986Sci...232..629S. doi:10.1126/science.232.4750.629. PMID 17781415.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ 67,0 67,1 Fastovsky, David E.; Sheehan, Peter M. (2005). "Reply to comment on "The Extinction of the dinosaurs in North America"" (PDF). GSA Today. 15 (7): 11. doi:10.1130/1052-5173(2005)015[11b:RTEOTD]2.0.CO;2. Geargiveer vanaf die oorspronklike (PDF) op 2 Junie 2018. Besoek op 21 Maart 2014.
- ↑ Sullivan, RM (2003). "No Paleocene dinosaurs in the San Juan Basin, New Mexico". Geological Society of America Abstracts with Programs. 35 (5): 15. Geargiveer vanaf die oorspronklike op 8 April 2011. Besoek op 2 Julie 2007.
- ↑ Fassett J.E., Heaman L.M., Simonetti A. (2011). "Direct U–Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico". Geology. 39 (2): 159–162. doi:10.1130/G31466.1.
{{cite journal}}: AS1-onderhoud: meer as een naam (link)
Verdere leesstof
[wysig | wysig bron]- Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction. New York: Morrow. ISBN 0-688-04287-2.
- Holtz, Thomas R. jr. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. New York: Random House. ISBN 978-0-375-82419-7.
- Paul, Gregory S. (2000). The Scientific American Book of Dinosaurs. New York: St. Martin's Press. ISBN 0-312-26226-4.
- Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: The Johns Hopkins University Press. ISBN 0-8018-6763-0.
- Sternberg, C. M. (1966). Canadian Dinosaurs, in Geological Series, no. 54. Tweede uitg. Ottawa: National Museum of Canada. p. 28
Eksterne skakels
[wysig | wysig bron] Wikimedia Commons het meer media in die kategorie Dinosourus.
Wikimedia Commons het meer media in die kategorie Dinosourus.
 Wikispecies het meer inligting verwant aan Dinosourus
Wikispecies het meer inligting verwant aan Dinosourus Hierdie artikel is vertaal uit die Engelse Wikipedia
Hierdie artikel is vertaal uit die Engelse Wikipedia